РАЗМНОЖЕНИЕ БАКТЕРИЙ И ПРИНЦИПЫ ИХ КУЛЬТИВИРОВАНИЯ
ЛФ и ПФ. Занятие № 4
| 7. Обмен веществ и энергии у прокариот | |
| 7.1. Особенности метаболизма микроорганизмов | |
| 7.2. Классификация бактерий по источнику углерода | |
| 7.3. Пути проникновения питательных веществ в бактериальную клетку | |
| 7.4. Классификация бактерий по потребностям в факторах роста | |
| 7.5. Классификация бактерий по особенностям энергетического метаболизма | |
| 7.6. Классификация бактерий по отношению к кислороду воздуха | |
| 7.7. Особенности метаболизма риккетсий, хламидий, микоплазм | |
| 7.8. Классификация бактериальных ферментов | |
| 8. Размножение бактерий и принципы их культивирования | |
| 8.1. Способы размножения бактерий | |
| 8.2. Цикл развития хламидий | |
| 8.3. Культивирование микроорганизмов и классификация искусственных питательных сред | |
| 8.4. Требования к условиям культивирования бактерий | |
| 8.5. Характер роста бактерий на искусственных питательных средах | |
| 8.6. Стадии роста периодической бактериальной культуры |
ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ У ПРОКАРИОТ
7.1. Особенности метаболизма микроорганизмов
Микроорганизмы обладают рядом принципиальных отличий в протекании метаболических процессов.
А. Бактерии, вследствие принципиально иной клеточной организации, по сравнению с эукариотическими клетками, обладают и принципиально иными чертами осуществления метаболических процессов, которые, в свою очередь, позволяют им занимать их уникальное положение в мире живых существ.
|
|
|
1. Бактерии, в целом, могут использовать любые источники химических соединений. Они «всеядны». Именно бактерии в круговороте веществ играют роль основного механизма минерализации органических веществ.
2. Прокариотическая клетка, по сравнению с эукариотической обладает несравненно более высокой скоростью метаболических процессов.
3. Бактерии в целом обладают высокой адаптационной способностью к меняющимся условиям окружающей среды. Эта способность реализуется у них как на клеточном, так и на популяционном уровнях.
а. На клеточном уровне эта способность детерминируется у прокариот высокой скоростью изменчивости; как фенотипической, так и генотипической.
б. Эта высокая изменчивость, реализуется, в свою очередь, в виде высокой гетерогенности бактериальной популяции. Поэтому, какие бы изменения во внешней среде не «обрушивались» на бактериальную популяцию, в ней всегда (как правило) найдется хотя бы одна клетка, которая, в силу своего отличия от других, сможет существовать в измененной внешней среде, и которая даст начало восстановлению прежней численности популяции. Именно этот процесс, прежде всего, имеют в виду, говоря об адаптации бактерий к меняющимся условиям окружающей среды.
|
|
|
Б. Вирусы, в отличие от клеточных форм жизни не обладают, строго говоря, собственным метаболизмом, используя метаболические потенции клетки-хозяина.
7.2. Классификация бактерий по источнику углерода
Так как жизнь на Земле основана на соединениях углерода, этот элемент является основным для построения и успешного функционирования бактериальной клетки.
А. Бактерии, использующие в качестве источника углерода неорганические соединения (углекислый газ, карбонаты), называются аутотрофами. Аутотрофы не вызывают патологических процессов в организме человека и не являются предметом изучения медицинской микробиологии.
Б. Медицинская микробиология имеет дело с бактериями, которые в качестве источника углерода используют органические соединения. Такие бактерии называются гетеротрофами.
1. Те гетеротрофы, которые могут использовать в качестве источника углерода так называемую «мертвую органику» (например, раствор глюкозы), называются сапрофитами.
|
|
|
2. Те гетеротрофы, которые используют в качестве источника углерода органические соединения живой метаболизирующей клетки, называются паразитами.
а. Подавляющее большинство бактерий этой группы могут использовать в качестве источника углерода как «мертвую», так и «живую» органику. Такие бактерии называются факультативными паразитами. Их можно культивировать на искусственных питательных средах, основанных на растворах органических соединений.
б. Риккетсии и хламидии не могут использовать в качестве источника углерода «мертвую» органику. Они могут размножаться только внутри живой клетки, используя ее органические вещества для своих метаболических процессов. Их называютоблигатными паразитами. Как и вирусы, эти бактерии не растут на искусственных питательных средах.
7.3. Пути проникновения питательных веществ в бактериальную клетку
Углеродсодержащие соединения, наравне с другими питательными веществами должны проникнуть в бактериальную клетку. Этот процесс аналогичен процессу питания более сложно организованных живых существ и поэтому иногда называется «питанием бактерий». У бактериальной клетки нет специализированных органелл для поглощения питательных веществ, последние проникают через клеточную оболочку по всему ее периметру. В большинстве случаев в этом процессе участвуют пермеазы – особые ферментоподобные белки. Пермеазы субстратспецифичны – для каждого вещества существует своя пермеаза, в обратимо связанном виде с которой оно перемещается через клеточную стенку и, если это требуется, цитоплазматическую мембрану.
|
|
|
А. Процесс проникновения питательных веществ через оболочку бактериальной клетки без затраты энергии называется диффузией. Энергия для осуществления диффузии не нужна, так концентрация диффундирующих веществ вне клетки выше, чем внутри ее – их перемещение происходит по градиенту концентрации.
1. Если в этом процессе не участвуют пермеазы, то это – простая диффузия. Таки путем внутрь бактериальной клетки проникают вещества, образующие истинные растворы.
2. Ряд субстратов (например, глицерин) диффундируют через оболочку бактериальной клетки с помощью пермеаз. Такой процесс называется облегченной диффузией.
Б. Концентрация подавляющего большинства необходимых питательных веществ внутри бактериальной клетки превышает их концентрацию в околоклеточном пространстве. Поэтому процесс их перемещения через клеточную оболочку происходит с затратой энергии. И, конечно, в этом процессе участвуют пермеазы.
1. Если при этом переносимые молекулы не претерпевают химических изменений, то такой процесс называется активным транспортом.
2. Если транспорт молекулы сквозь клеточную оболочку сопровождается ее химической модификацией (например, фосфорилированием), то такой процесс называется транслокацией химических групп.
7.4. Классификация бактерий по потребностям в факторах роста
Факторами роста (или ростовыми факторами) понимают макромолекулы (азотистые основания, аминокислоты, витамины, липиды и пр.), необходимые для построения и функционирования бактериальной клетки.
А. Бактерии, способные самостоятельно синтезировать все необходимые им факторы роста из более простых веществ (в типичном случае – солей аммония и глюкозы), называются прототрофами.
Б. Ауксотрофы не способны синтезировать один или несколько факторов роста. Чтобы выращивать такие бактерии на искусственной питательной среде, необходимо, чтобы эта среда содержала нужные ростовые факторы «в готовом виде».
7.5. Классификация бактерий по особенностям энергетического метаболизма
Этот процесс аналогичен процессу дыхания более сложно организованных живых существ и поэтому иногда называется «дыханием бактерий».
А. Бактерии, которые получают энергию непосредственно солнечного света, называются фототрофами. Такие бактерии не вызывают патологические процессы в организме человека и медицинской микробиологией не изучаются.
Б. Бактерии, которые получают энергию, разлагая более сложные химические вещества до более простых в процессе окислительно-восстановительных реакций с запасанием полученной энергии в виде АТФ и других макроэргических соединений, называются хемотрофами.
1. Донорами электронов при этом могут выступать как неорганические, так и органические соединения.
а. Хемотрофы, использующие в качестве доноров электронов в химических реакциях, сопровождающихся синтезом АТФ, неорганические соединения, называются литотрофами.
б. Хемотрофы, использующие в качестве доноров электронов в химических реакциях, сопровождающихся синтезом АТФ, органические соединения, называются органотрофами.
2. Акцептором электронов в таких реакциях может служить молекула, которая находится или вне бактериальной клетки (так называемый внешний акцептор электронов) или внутри бактериальной клетки (в этом случае говорят о внутреннем акцепторе электронов).
а. Цепь окислительно-восстановительных реакций для получения макроэргических соединений с переносом электронов на внешний акцептор называется окислением. Именно для обозначения этого процесса в бактериологии используется термин «дыхание».
1. Если в качестве такого внешнего акцептора электронов выступает молекулярный кислород, то такой процесс называется аэробным дыханием.
2. В качестве внешнего акцептора электронов некоторые бактерии (например, клостридии) используют не молекулярный кислород, а нитрат или фумарат. В этом случае используют термин анаэробное дыхание (нитратное или фумаратное дыхание).
б. Цепь окислительно-восстановительных реакций для получения макроэргических соединений с переносом электронов на внутренний акцептор называется ферментацией или брожением.
7.6. Классификация бактерий по отношению к кислороду воздуха
Вовлечение кислорода воздуха в окислительно-восстановительные реакции сопровождается образованием перекисного радикала (О•) – атома кислорода с лишним электроном на внешней оболочке. Это очень сильный окислитель. В зависимости от того, как бактерии решают проблему его нейтрализации, и зависит их способность существовать в кислородной среде. На практике простейшим методом определить отношение изучаемого вида бактерий к кислороду воздуха, является регистрация характера роста этих бактерий при засеве уколом в столбик полужидкой среды (Рис. 7-1).
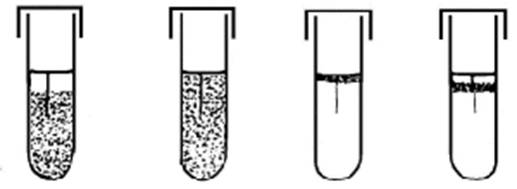 Рис. 7-1. Рост (слева направо) облигатных анаэробов, факультативных анаэробов, облигатных аэробов и микроаэрофилов при засеве уколом в столбик полужидкого агара
Рис. 7-1. Рост (слева направо) облигатных анаэробов, факультативных анаэробов, облигатных аэробов и микроаэрофилов при засеве уколом в столбик полужидкого агара
|
А. Бактерии, которым кислород абсолютно необходим как внешний акцептор электронов, образовавшийся перекисный радикал с помощью фермента супероксиддисмутазы переводят в перекись кислорода. Это соединение также является окислителем, хотя и менее сильным, чем перекисный радикал. Перекись водорода нейтрализуется с образованием воды и молекулярного кислорода с помощью, например, каталазы. В отсутствии кислорода такие бактерии не растут, так как не могут использовать другие молекулы в качестве акцептора электронов, и поэтому называются облигатными аэробами.
Б. Размножение некоторых бактерий возможно только в атмосфере особого газового состава. Такие бактерии обладают супероксиддисмутазой, но наличие каталазы или других ферментов, нейтрализующих перекись водорода, может быть вариабельно.
1. Микроаэрофилы растут при более низком, чем в атмосферном воздухе, содержании кислорода. К ним относятся, например, молочнокислые бактерии, в большом количестве населяющие кишечник человека.
2. Капнофилы нуждаются в более высоком, чем в атмосферном воздухе, содержании углекислого газа. К ним, например, относятся бруцеллы, вызывающие бруцеллез.
3. Существуют также бактерии, которые требуют для своего успешного культивирования создания особой газовой смеси, включающей как пониженное количество кислорода, так и повышенное – углекислого газа. Т.е. такие бактерии можно определить одновременно как микроаэрофилы и капнофилы. Такие особые требования к аэрации при культивировании предъявляют, например, кампилобактеры и геликобактер.
В. Подавляющие большинство бактерии могут расти в отсутствии кислорода, вследствие чего их называют анаэробами.
1. Некоторые бактерии лишены не только ферментов, нейтрализующих перекись водорода, но и супероксиддисмутазы. В результате чего они погибают при малейшем контакте с атмосферным воздухом и называются облигатными анаэробами. При микробиологической диагностике болезней, вызываемых этой группой бактерий, и взятие патологического материала и посев его на питательные среды, не говоря уже о самом культивировании, необходимо проводить с соблюдением специальных приемов, исключающих контакт возбудителя с атмосферным воздухом.
2. Часть анаэробов, не имея ферментов, нейтрализующих перекись водорода, содержат супероксиддисмутазу. Такие бактерии – аэротолерантные анаэробы – выдерживают кратковременный контакт с атмосферным воздухом, например во время взятия патологического материала или засева его на питательную среду, но культивироваться могут лишь в анаэробных условиях.
3. Однако наиболее часто медицинская микробиология имеет дело с бактериями, которые имеют полный набор ферментов, позволяющих им, расти и размножаться и в присутствии и в отсутствии кислорода воздуха – т.е. в любых условиях аэрации. Такие бактерии называются факультативными анаэробами.
7.7. Особенности метаболизма риккетсий, хламидий, микоплазм
Риккетсии, хламидии, а также микоплазмы имеют, по сравнению с остальными прокариотами, принципиальные отличия метаболических процессов.
А. Риккетсии не способны синтезировать некоторые макромолекулы, вследствие чего вынуждены получать их в готовом виде от клетки-хозяина. Поэтому они могут функционировать только внутри живой эукариотической клетки (облигатный внутриклеточный паразитизм).
Б. Хламидии по той же причине, что и риккетсии, являются облигатными внутриклеточными паразитами, но, кроме этого они еще и– их часто называют «энергетическими паразитами». не способны самостоятельно синтезировать АТФ
В. Патогенные микоплазмы не способны синтезировать стерины для своей цитопалзматической мембраны и «высасывают» их из клеточной мембраны клетки-хозяина, что дало повод назвать их «мембранными паразитами».
7.8. Классификация бактериальных ферментов
Ферменты бактериальной клетки классифицируются так же, как и ферменты эукариот – в зависимости от типа биохимических реакций, в которых они участвуют. Однако, в бактериологии используется и классификация бактериальных ферментов в зависимости от их локализации, зависимости их синтеза от наличия субстрата; кроме этого отдельную группу составляют ферменты вирулентности (или ферменты патогенности – различия между терминами «патогенность» и «вирулентность» будут рассмотрены ниже).
А. По своей локализации бактериальные ферменты подразделяются на две группы.
1. Ферменты, которые синтезируются бактериальной клеткой в окружающую среду и действуют в околоклеточном пространстве, называются экзоферментами. К ним относятся, например, протеолитические ферменты грамположительных бактерий, пеницилиназы и т.п. Такая локализация ферментов обусловлена их функциональной задачей – они, например, расщепляют макромолекулярный питательный субстрат на низкомолекулярные части, способные транспортироваться внутрь клетки, или инактивируют вещества, обладающие антибактериальной активностью, «на подходе» к клетке.
2. Ферменты, которые локализуются внутри бактериальной клетки, называются эндоферментами. Эндоферменты содержатся в периплазматическом пространстве грамотрицательных бактерий (здесь находится часть тех ферментов, которые у грамположительных бактерий являются экзоферментами, например, протеазы), а также в цитоплазматической мембране и цитоплазме.
Б. По зависимости своего синтеза от наличия субстрата, на который действует фермент, бактериальные ферменты подразделяются также на две группы.
1. Конститутивные ферменты синтезируются постоянно, вне зависимости от наличия в данный момент субстрата для их действия.
2. Индуцибельные ферменты синтезируются бактериальной клеткой только если «в пределах досягаемости» находится субстрат для их действия. Типичным примером этой группы ферментов являются ферменты лактозного оперона.
В. К ферментам вирулентности относятся такие бактериальные ферменты, чьим субстратом действия являются клетки и ткани макроорганизма, в котором паразитирует микроб. Более подробно эта группа бактериальных ферментов будет охарактеризована в 19 главе.
РАЗМНОЖЕНИЕ БАКТЕРИЙ И ПРИНЦИПЫ ИХ КУЛЬТИВИРОВАНИЯ
8.1. Способы размножения бактерий
Термины «размножение бактерий» и «рост бактерий» часто используют как синонимы, хотя эти термины обозначают, строго говоря, разные явления. Под ростом бактерий понимают увеличение размеров бактериальной клетки, а под размножением бактерий – увеличение числа бактериальных клеток. Однако когда имеют в виду бактериальную популяцию в целом, то в этом случае термином «рост» обозначают увеличение количества особей в популяции. В последнем случае корректней использовать термин «рост бактериальной культуры».
А. Основной способ размножения для большинства бактерий – бинарное деление.
1. У грамположительных бактерий бинарное деление происходит путем формирования перегородки от противоположенных концов клеточной стенки к центру, где обе части перегородки сливаются, сформировав тем самым две самостоятельные клетки.
2. У грамотрицательных бактерий бинарное деление происходит путем образования перетяжки: клетка как бы истончается посередине, пока не разорвется на две самостоятельные клетки.
Б. Ряд бактерий могут делиться путем почкования (например, франциселлы, микоплазмы).
В. Те бактерии, которые формируют нитевидные формы, могут делиться путем их фрагментации (например, актиномицеты, микоплазмы).
Г. У стрептомицетов существует способ размножения экзоспорами.
Д. У хламидий существует особый цикл развития (см. ниже).
8.2. Цикл развития хламидий
Хламидии являются облигатными внутриклеточными паразитами и могут размножаться только внутри эукариотической клетки. Но при этом они должны обладать способностью сохраняться во внеклеточной среде, чтобы после выхода из своей клетки-хозяина найти и поразить следующую. У хламидий для этого существуют различные клеточные формы (Рис. 8-1 и 8-2).
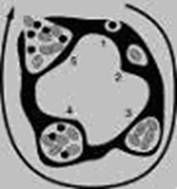 Рис. 8-1. Общая схема жизненного цикла хламидий
Рис. 8-1. Общая схема жизненного цикла хламидий
|  Рис. 8-2. Электронограмма клетки, пораженной хламидиями
(N – ядро, eb – элементарное тельце, rb – ретикулярное тельце)
Рис. 8-2. Электронограмма клетки, пораженной хламидиями
(N – ядро, eb – элементарное тельце, rb – ретикулярное тельце)
|
А. Элементарное тельце выполняет инфекционную функцию – проникает в клетку-хозяина путем инвагинации места адсорбции.
Б. Ретикулярное (или инициальное) тельце размножается бинарным делением в образовавшемся цитоплазматическом пузырьке и формирует микроколонию, которая видна при микроскопии как цитоплазматическое включение.
В. Переходную форму от ретикулярного тельца к тельцу элементарному представляет промежуточное тельце. После того, как в микроколонии сформировалось множество новых элементарных телец, она сливается с клеточной мембраной и «изливается» наружу «урожаем» размножившихся хламидий, элементарные тельца которых отправляются на поиски новых клеток-хозяев.
8.3. Культивирование микроорганизмов и классификация искусственных питательных сред
Есть два основных принципа культивирования микроорганизмов – in vivo и in vitro.
А. Микроорганизмы – облигатные паразиты (риккетсии, хламидии, вирусы) можно культивировать только in vivo, т.е. внутриклеточно. Для этого используют методы, разработанные вирусологами и разбираемые в курсе вирусологии – культивирование микроорганизмов в культуре клеток, в птичьем эмбрионе, в организме лабораторного животного.
Б. Микроорганизмы – факультативные паразиты (т.е. все бактерии, кроме риккетсий и хламидий) можно культивировать in vitro – на искусственных питательных средах, которые классифицируются по своей консистенции, составу и назначению.
1. В зависимости от их консистенции искусственные питательные среды классифицируются на плотные, полужидкие и жидкие (Рис. 8-3).
а. Плотные питательные среды могут быть агаризованные и свернутые.
1. Агаризованные питательные среды называются так потому, что в качестве уплотнителя в их состав вводят агар – полисахарид, добываемый из морских водорослей определенных видов и используемый для уплотнения питательных сред в бактериологии по такому же алгоритму, как в быту крахмал или желатин. Для получения плотной питательной среды достаточно концентрации агара в пределах 1,5 – 2%. При этом могут использоваться или чашки Петри («пластинчатый агар») или пробирки (при наличии скоса –«скошенный агар» или «косяк», при его отсутствии – «столбик агара»).
 Рис. 8-3. Питательные среды для культивирования бактерий
(слева направо – пластинчатый агар, столбик, бульон, косяк) Рис. 8-3. Питательные среды для культивирования бактерий
(слева направо – пластинчатый агар, столбик, бульон, косяк)
|
2. Свернутые питательные среды – это плотные среды, содержащие сыворотку крови или обогащенные другим белком (яичные, например), которые уплотняются (как результат денатурации белка) при их прогревании в процессе стерилизации.
б. Полужидкие питательные среды содержат небольшое количество агара (примерно 0,5%).
в. Жидкие питательные среды не содержат уплотнителей. В принципе, любую жидкую среду можно превратить в плотную, или добавив к ней агар, или свернув содержащийся в ней белок (соблюдая в обоих случаях необходимую концентрацию уплотнителя).
2. По своему составу искусственные питательные среды классифицируются на натуральные и синтетические.
а. Натуральные искусственные питательные среды готовятся на основе отваров или экстрактов мяса, рыбы, овощей и др. натуральных продуктов. Натуральные питательные среды, свою очередь, классифицируют на простые и сложные. Именно преимущественно натуральные питательные среды используются в работе бактериологических лабораторий так называемого «практического здравоохранения».
1. Простые натуральные питательные среды, собственно, и представляют собой такие отвары или экстракты. К простым натуральным питательным средам относятся:
– мясопептонный агар (МПА) и мясопептонный бульон (МПБ), относящиеся к одной группе, так как агар не усваивается подавляющим большинством бактерий, и, следовательно, он не изменяет состав среды – только ее концентрацию;
– желатин;
– молоко;
– кусочки овощей.
2. Сложные натуральные питательные среды получают путем добавления в простые натуральные среды любого вещества (красителя, сахара, антибиотика, крови и т.д.).
б. Синтетические искусственные питательные среды получают, смешивая в растворе чистые химические вещества (как правило, соли). В отличие от натуральных питательных сред, это так называемые среды известного состава, так как количество содержащихся в них веществ точно задается рецептурой их приготовления. Синтетические искусственные питательные среды используются в основном в бактериологических лабораториях научных учреждений.
3. По своему назначению искусственные питательные среды классифицируются на основные, элективные (селективные), дифференциально-диагностические и консервирующие.
а. Основные искусственные питательные среды названы так потому, что с их помощью проводится основная работа бактериолога – накопление чистой культуры, ее «оживление» после длительного хранения и т.п. Основные питательные среды, в свою очередь, подразделяются на универсальные и специальные.
1. Одна и та же универсальная основная питательная среда может быть использована для культивирования многих видов бактерий. По своему составу это – простые натуральные питательные среды. Бактерии, которые можно культивировать на таких средах, называются бактериями с простыми питательными потребностями.
2. Специальная основная питательная среда используется для культивирования конкретного вида или группы бактерий. Бактерии, которые для своего культивирования нуждаются в специальных питательных средах, называются бактериями со сложными питательными потребностями.
б. Элективные (селективные, избирательные, обогащения) искусственные питательные среды – это среды, содержащие вещества, используемые бактериями определенных видов и не благоприятствующие или даже препятствующие росту других бактерий. Такие среды служат для выделения конкретного вида бактерий из патологического материала. Выделение тех виды бактерий, для которых такие среды не разработаны, довольно затруднительно.
в. Дифференциально-диагностические искусственные питательные среды – это среды, позволяющие отличать одни виды бактерий от других по их ферментативной активности (чаще всего – по цвету образуемых ими колоний) или культуральным свойствам.
г. Консервирующие искусственные питательные среды – это среды, используемые, например, при доставке патологического материала в бактериологическую лабораторию; так как метаболическая активность на них бактерий сводится практически к нулю, то бактерии сохраняются, но не размножаются.
8.4. Требования к условиям культивирования бактерий
Для, того, чтобы успешно культивировать бактерии на искусственных питательных средах, необходимо учитывать не только их питательные потребности (простые или сложные, о чем было сказано выше), но и температуру культивирования, реакцию среды (рН), а также необходимые конкретному виду условия аэрации.
А. По оптимальной температуре культивирования бактерии классифицируются на три группы: термофилы, мезофилы и психрофилы.
1. Оптимальная температура культивирования термофилов составляет 50 – 60°С. По понятным причинам термофилы не составляют предмет изучения медицинской микробиологии.
2. Подавляющее большинство бактерий, имеющих медицинское значение, лучше всего растут при температуре человеческого организма, т.е. 37°С. Такие бактерии называются мезофилами.
3. Ряд патогенных для человека бактерий лучше всего растут при более низких температурах (от 6 до 20°С) и называются психрофилами.
Б. В зависимости от необходимой реакции питательной среды бактерии подразделяются на три основные группы, для обозначения двух из которых используются особые термины.
1. Те из них, которые лучше всего растут на кислых питательных средах, называются ацидофилами.
2. Бактерии, для культивирования которых необходимо использовать щелочные среды, называются алкалифилами.
3. Подавляющее большинство патогенных для человека бактерий растут на средах с нейтральным рН. Для обозначения этой группы бактерий никаких особых терминов не используется.
В. По требованиям к условиям аэрации во время культивирования, бактерии можно разделить на четыре основные группы.
1. Облигатные аэробы требуют во время культивирования постоянного доступа воздуха к поверхности питательной среды.
2. Анаэробы, наоборот, культивируются в безвоздушной среде.
3. Особого газового состава для своего культивирования требуют микрофилы (сниженного содержания кислорода) и капнофилы (повышенного содержания углекислого газа).
4. Факультативные анаэробы растут при любых условиях аэрации.
8.5. Характер роста бактерий на искусственных питательных средах
Характер роста бактерий зависит, прежде всего, от того, какая питательная среда – жидкая или плотная – используется для культивирования.
А. На жидких питательных средах (например, мясопептонном бульоне) для всего многообразия бактерий можно выделить четыре формы роста.
1. Большинство бактерий формируют диффузную муть (Рис. 8-4).
2. Так называемые «коховские бактерии», названные так потому, что их основные патогенные представители были открыты Кохом, – бациллы, микобактерии и вибрионы – образуют на поверхности бульона пленку.
3. Для стрептококков характерен так называемый придонный или пристеночный рост – осадок или мелкие хлопья у стенки пробирки при прозрачном бульоне.
4. Возбудитель чумы – Yersinia pestis – растет в виде пленки на поверхности бульона, от которой спускаются тяжи, похожие на сталактиты, может формироваться и осадок.
Б. На плотных питательных средах (например, мясопептонном агаре) рост бактерий зависит от способа их засева.
1. Если плотность засева большая, то бактерии формируют на поверхности агара сплошной налет – так называемый, «рост газоном» или «сливной рост».
2. Если засев проводится таким образом, что каждая бактериальная клетка лежит на поверхности агара на большом расстоянии от других, то, после многократных делений она формирует изолированную колонию (говорят еще об «изолированном росте»). А так, как колония – результат размножения одной клетки, то ее, с некоторыми допущениями, рассматривают как клональную культуру. Именно из материала отдельной, изолированной, колонии в процессе культурального метода исследования получают так называемую «чистую культуру» – культуру, содержащие клетки только одного вида. Все огромное многообразие, по их внешнему виду, колоний (Рис. 8-5) можно свести к двум основным типам.
 Рис. 8-4. Рост бактерий на МПБ в виде диффузной мути
(сравните с видом чистого МПБ на предыдущем рисунке) Рис. 8-4. Рост бактерий на МПБ в виде диффузной мути
(сравните с видом чистого МПБ на предыдущем рисунке)
|
 Рис. 8-5. Колонии бактерий на пластинчатом агаре Рис. 8-5. Колонии бактерий на пластинчатом агаре
|
а. S-форма колонии («гладкая») – гомогенная, с ровными краями, куполообразная, влажная, прозрачная или полупрозрачная. Все S-форма колоний схожи друг с другом, отличаясь у разных видов бактерий или их вариантов размером, а в случае пигментообразования или роста на дифференциально-диагностических средах – и цветом. S-форму колоний образуют:
– кокки,
– грамотрицательные палочки, кроме возбудителя чумы (Yersinia pestis).
б. R-форма колоний («шероховатая») – не гомогенная, с неровными краями, с самыми разнообразными вариантами расположения относительно поверхности питательной среды (от возвышающейся над ней до погруженной в нее, т.е. находящейся ниже поверхности питательной среды), непрозрачная. R-формы колоний различных бактерий могут резко отличаться друг от друга, для их описания порой используют сравнительный обороты (говорят, например, о колониях, похожих на цветок маргаритки у возбудителя дифтерии, похожих на цветную капусту или бородавку – у возбудителей туберкулеза и т.п.). R-форма колоний образуют:
– грамположительные палочки,
– возбудитель чумы (Yersinia pestis).
8.6. Стадии роста периодической бактериальной культуры
При выращивании бактерий в жидкой питательной среде можно постоянно отбирать выросшую бактериальную массу, удалять продукты метаболизма бактерий и добавлять новую полноценную питательную среду. Т.е. постоянно поддерживать оптимальные условия культивирования. В этом случае культура будет постоянно расти с максимальной скоростью. Такая культура называется хемостатной, потому что для ее получения используют специальные приборы - хемостаты, – которые и дают возможность совершать вышеописанные манипуляции. Такие культуры используются в промышленной микробиологии для получения полезных веществ – продуктов микробного метаболизма (антибиотиков, аминокислот и т.д.). Если же бактериальная культура выращивается в пробирке (именно такой способ используется в медицинской микробиологии), то с течением времени в питательной среде накапливаются продукты бактериального метаболизма, а питательная среда, наоборот, истощается. В результате, чтобы культура не погибла, ее необходимо периодически пересевать на свежую питательную среду. Такая культура называется периодической. В своем росте она проходит девять стадий (фаз) развития.
А. После внесения в питательную среду инокулята (посевной дозы), наступает лаг-фаза (Рис. 8-6). Деление клеток на этой стадии не происходит – бактерии как бы приспосабливаются к новой среде обитания; при этом некоторое количество их может погибнуть.
Б. Затем наступает фаза положительного ускорения (Рис. 8-7). Бактериальные клетки начинают делиться и скорость их деления постоянно увеличивается.
 Рис. 8-6. Стадии роста периодической бактериальной культуры: лаг-фаза
Рис. 8-6. Стадии роста периодической бактериальной культуры: лаг-фаза
|  Рис. 8-7. Стадии роста периодической бактериальной культуры: фаза положительного ускорения
Рис. 8-7. Стадии роста периодической бактериальной культуры: фаза положительного ускорения
|
В. Наконец, скорость деления клеток достигнет максимального значения и останется таковой некоторое время (Рис. 8-8). Это экспоненциальная фаза (фаза логарифмического роста).
Г. Однако, со временем количество питательных веществ в среде снижается, а концентрация продуктов метаболизма бактериальных клеток увеличивается. В результате условия для размножения и роста бактерий ухудшаются, и скорость деления бактериальных клеток снижается – наступает фаза отрицательного ускорения (Рис. 8-9). Однако, на этой стадии количество живых бактериальных клеток в культуре все еще увеличивается, правда с постоянно снижающейся скоростью.
 Рис. 8-8. Стадии роста периодической бактериальной культуры: фаза логарифмического роста (экспоненциальная)
Рис. 8-8. Стадии роста периодической бактериальной культуры: фаза логарифмического роста (экспоненциальная)
|  Рис. 8-9. Стадии роста периодической бактериальной культуры: фаза отрицательного ускорения
Рис. 8-9. Стадии роста периодической бактериальной культуры: фаза отрицательного ускорения
|
Д. Стадия, во время которой в каждый конкретный момент времени количество вновь появившихся живых бактериальных клеток равно количеству погибших, называется стационарной фазой максимума (Рис. 8-10). Бактериальная культура, достигшая в этой фазе роста максимально возможной при данных условиях культивирования концентрации живых бактериальных клеток, называют остановившейся.
Е. Затем количество живых бактериальных клеток начинает уменьшаться с увеличивающейся скоростью. Эта стадия роста бактериальной культуры называется фазой ускоренной гибели (Рис. 8-11).
 Рис. 8-10. Стадии роста периодической бактериальной культуры: стационарная фаза максимума
Рис. 8-10. Стадии роста периодической бактериальной культуры: стационарная фаза максимума
|  Рис. 8-11. Стадии роста периодической бактериальной культуры: фаза ускоренной гибели
Рис. 8-11. Стадии роста периодической бактериальной культуры: фаза ускоренной гибели
|
Ж. Стадия, во время которой эта скорость убывания живых бактериальных клеток становится максимальной, называется фазой логарифмической гибели (Рис. 8-12).
З. Со временем, однако, эта скорость начинает уменьшаться – наступает фаза уменьшения скорости гибели (Рис. 8-13).
 Рис. 8-12. Стадии роста периодической бактериальной культуры: фаза логарифмической гибели
Рис. 8-12. Стадии роста периодической бактериальной культуры: фаза логарифмической гибели
|  Рис. 8-13. Стадии роста периодической бактериальной культуры: фаза уменьшения скорости гибели
Рис. 8-13. Стадии роста периодической бактериальной культуры: фаза уменьшения скорости гибели
|
И. В конце концов культура некоторое время будет состоять из минимального количества живых бактериальных клеток. Эта стадия роста бактериальной культуры называется стационарной фазой минимума (Рис. 8-14). Время, которое бактериальная культура сможет продержаться до своей гибели на этот минимуме, зависит как от вида микроорганизмов, так и от условий культивирования.
| Рис. 8-14. Стадии роста периодической бактериальной культуры: стационарная фаза минимума |
Дата добавления: 2018-05-02; просмотров: 268; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!