Развитие потенциала действия.
Лекция 11 Физика нервного импульса (потенциала действия)
Биолектрические явления, рассмотренные в предыдущих лекциях, играют ключевую роль в процессах распространения нервного импульса, обеспечивающих передачу информации от центральной нервной системы к периферической (моторные, или эфферентные сигналы) и обратную передачу (сенсорные, или афферентные сообщения) (Антонов В.Ф. Физика и биофизика 2012. С. 245…. Рубин А.Б. Биофизика т 2 М. 2000 стр 166….).
Посредством электрических нервных импульсов (потенциалов действия) в живом организме передается информация от рецепторов к нейронам мозга и от нейронов мозга к мышцам. Живой организм является полностью электрифицированной системой. Без электричества нет
Потенциал действия был открыт раньше потенциала покоя. Животное электричество известно давно. Разряды электрического угря (происходящие при напряжении до 600 В, с током около 60 А и длительностью порядка миллисекунды) использовались медициной еще в Древнем Риме для лечения подагры, головной боли, эпилепсии. Электрический нервный импульс открыл Луиджи Гальвани, профессор анатомии в г. Болонья. Результаты его электрофизиологических опытов изложены в книге "Трактат о силах электричества при мышечном движении" (1791 г.). Гальвани открыл, что мышечные сокращения конечностей препарированной лягушки могут вызваться электрическим импульсом. Однако,
То, что нервный импульс представляет собой импульс электрического тока, было доказано лишь к середине 20-го века, в основном в работах английского физиолога А. Ходжкина и его сотрудников. В 1963 году Ходжкину, Хаксли и Иклсу была присуждена Нобелевская премия по медицине "за оперирование нервных клеток " (аксон кальмара).
Строение нейрона.
Как известно, функциональной единицей нервной системы является нейрон (см. рис 1) .

Рис.1
Особенностью данной клетки является наличие длинного отростка –аксона, по которому и распространяется нервный импульс. Аксон частично может быть покрыт миелиновой оболочкой ((многократно (200 раз) закрученные мембраны Шванновских клеток), играющей роль электроизолятора между мембраной аксона и окружающим электролитом. Неизолированные участки называются перехватами Ранвье. Аксон заканчивается нервными окончаниями – синапсами. Кроме Аксона имеются и короткие отростки – дендриты, также оканчивающиеся синапсами и обеспечивающие обмен информацией между различными нейронами. Размеры клеточного тела нейрона, его ядра и дентритов типичны для всех клеток (1…100мкм). В то же время длина аксона может достигать нескольких метров при диаметре до сотен микрометров. Аксоны группируются в пучки – нервы. Каждый нерв состоит из большого числа аксонов разного диаметра, обеспечивающих передачу информации в нужном направлении.
Потенциал действия
Если в каком либо месте локально возбудить аксон, то по нему начнет распространяться отдельный электрический импульс – потенциал действия или спайк.
При этом величина, форма, и длительность отдельного импульса не зависят от природы и силы раздражителя.
Аксоны могут возбуждаться электрическими импульсами различной формы, теплом, холодом, химическим воздействием и механическим напряжением – все это приводит к одинаковому эффекту. Скорость распространения нервного импульса была измерена уже Гельмгольцем. При сравнимых диаметрах аксонов она больше у миелинизированных волокон (у кошки: 0.7…2.3 м/с –немиелинизированные волокна, 10…100м/с - миелинизированные волокна).
В лабораториях потенциал действия обычно возбуждают с помощью электрического тока, так как при этом легко контролировать силу, время и длительность раздражителя. Типичная схема эксперимента показана на рис.2 (слева).
Процесс нервного возбуждения развивается следующим образом Если пропустить через аксон слабый импульс тока Io, приводящий к деполяризации мембраны, то после снятия внеш. воздействия Io (1a) потенциал  (1б) монотонно возвращается к исходному уровню. В этих условиях аксон ведёт себя как пассивная электрич. цепь, состоящая из конденсатора и пост. сопротивления.
(1б) монотонно возвращается к исходному уровню. В этих условиях аксон ведёт себя как пассивная электрич. цепь, состоящая из конденсатора и пост. сопротивления.

Рис. 2. Развитие потенциала действия в нервном волокне:а- подпороговое (1) и надпороговое (2)раздражения;б-мембранный отклик; при надпороговом раздражении проявляется полный потенциал действия 2б;в- ионный ток, протекающий черезмембрану при возбуждении;г- аппроксимацияионного тока в простой аналитической модели.
Если импульс тока превышает нек-рую пороговую величину, потенциал продолжает изменяться и после выключения возмущения; потенциал становится положительным и только потом возвращается к уровню покоя, причём вначале даже несколько проскакивает его (область гиперполяризации, рис. 2). Отклик мембраны при этом не зависит от возмущения; этот импульс наз. потенциалом действия. Одновременно через мембрану течёт ионный ток, направленный сначала внутрь, а потом наружу (рис. 2,в).
Классические эксперименты такого рода были выполнены Ходжкиным и Хаксли (1952г).Они исследовали распространение нервного импульса в гигантских аксонах кальмаров (диаметр -600 мкм). Для возбуждения потенциала действия подавалось деполяризирующее напряжение (полярность напряжения противоположна полярности потенциала покоя).
В допороговой области (Io ≤ Ic) данное воздействие приводило лишь к смещению исходного отрицательного потенциала. При превышении порога – наблюдалась генерация потенциала действия - быстрая деполяризация мембраны (изменение знака потенциала) с последующей реполяризацией. Зависимость потенциала действия от времени имела сложный и немонотонный характер. Потенциалы, регистрируемые в различных точках имели один и тот же вид, но были сдвинуты по времени
Измерения показали, что пороговый ток возбуждения Ic существенно зависит от длительности импульса. При очень больших длительностях этот ток минимален, но не равен нулю. Это минимальное значение тока Imin называется реобазой. При уменьшении длительности импульсов значение порогового тока возрасатает, причем в области малых длительностей порог постоянен по заряду (qmin =Ic* Δt = const) , а не по току. Отношение порогового заряда к реобазе
Δtхр = qmin/ Imin соответствует переходу от одного типа поведения потенциала к другому типу поведения.
Если сообщить аксону два последовательных импульса возбуждения, то его отклик зависит от интервала времени, разделяющего эти импульсы. При малом интервале времени второй импульс приходит в момент времени, в которой аксон еще возбужден предыдущим импульсом, т.е. находится в абсолютном рефрактерном состоянии и не может возбудиться снова. Затем наступает относительное рефрактерное состояние, когда для возбуждения аксона требуются большие амплитуды импульса. Продолжительность рефрактерного периода – несколько миллисекунд.
Приведенные экспериментальные данные свидетельствуют о сложных механизмах, лежащих в основе распространения нервного импульса. Действительно, прямое кабельное распространение невозможно из-за высокого электрического сопротивления аксоплазмы .
Задача:Рассчитать сопротивление участка гигантского аксона кальмара с диаметром 600мкм и длиной 1м используя максимальные значения удельной электропроводности аксоплазмы – 10 Ом.м.
Ответ: около 300 Мом.
При столь высоких сопротивлениях электрический сигнал должен сильно затухать, а экспериментально было установлено, что потенциал действия распространяется практически без затухания.
Действительный механизм распространении нервного импульса был предложен Ходжкиным и Хаксли. Они обратили внимание на то, что при удалении из внеклеточной среды ионов натрия потенциал действия не возникает. Таким образом, его появление было объяснено резким возрастанием проводимости мембраны относительно ионов натрия. Это подтверждается и тем, что на пике потенциала действия его значение близко к равновесному значению для ионов натрия.
Ионные каналы. В мембранах нервной клетки имеются каналы, проницаемые для ионов Na+ , K+, Са2+ и Сl-. Эти каналы чаще всего находятся в закрытом состоянии и открываются лишь на короткое время. Каналы подразделяются на потенциал-управляемые (илиэлектровозбудимые), например быстрые Na+ каналы, и лиганд-управляемые (или хемовозбудимые). Каналы — это интегральные мембранные белки, состоящие из многих субъединиц. В зависимости от изменения мембранного потенциала или взаимодействия с соответствующими лигандами, нейромедиаторами и нейромодуляторами, белки-рецепторы могут находиться в одном их двух конформационных состояний, что и определяет проницаемость канала («открыт» — "закрыт» —и т.д.).
Развитие потенциала действия.
1. По мере приближения мембранного потенциала действия к пороговому значению открываются натриевые каналы – возникает входящий ток, переносимый ионами Na+. В допороговой области приток ионов натрия компенсируется оттоком ионов калия через калиевые каналы.
2.При превышении порога данная компенсация уже невозможна. Это связано с различной зависимостью проницаемости натриевого и калиевого каналов от напряжения. Na- проводимость быстро лавинообразно возрастает (открываются новые Na- каналы) и столь же быстро уменьшается по экспоненте К – проводимость по S-образной кривой и за 5-6 мс выходит на постоянный уровень. Восстановление Na- проводимости до исходного значения происходит в 10 раз быстрее чем К-проводимости.
3. В результате сильного потока ионов натрия происходит деполяризация мембраны. При приближении потенциала действия к пику, э.д.с. = (Vm – εNa), действующая на ионы натрия уменьшается, скорость изменения потенциала тоже уменьшается – достигается пик.
4. Начинается инактивация – закрытие Na каналов, что приводит к реполяризации мембраны, которая усиливается за счет задержки открывания потенциал управляемых калиевых каналов.
5. Начинается открывание К+ каналов и так как на ионы калия действует ЭДС =(Vm – εк), из клетки быстро выходит небольшое количество ионов калия и мембрана реполяризуется до состояния покоя.
Инактивация Na+ каналов и высокая К+ проводимость сохраняется после завершения потенциала действия (ПД) в течении нескольких мс. Во время абсолютного рефрактерного периода открытых натриевых каналов недостаточно для компенсации выхода ионов калия. Во время относительного рефрактерного периода сигнал большой амплитуды может активировать большое число натриевых каналов и вызвать потенциал действия.
Указанными механизмами можно объяснить явление аккомодации. Разные мембраны имеют разную степень аккомодации, это связано с соотношением натриевых и калиевых каналов.
Важно подчеркнуть, что количество ионов, проходящих через мембрану при одном потенциале действия (ПД) относительно мало и практически не вызывает измения внутриклеточной концентрации ионов (исключение – мельчайшие нервные клетки. Для возникновения ПД величиной 100 мВ необходимо переместить 160 ионов натрия через 1мкм2 (на самом деле – 500, т.к. вход натрия частично компенсируется выходом калия. При этом концентрация ионов внутри гигантского аксона кальмара меняется на 0.001%. Поэтому если натриевый насос этого аксона вывести из строя метаболическим ядом он еще способен сгенерировать несколько тысяч импульсов. Для тонких аксонов с диаметром 1мкм при каждом ПД концентрация изменяется на 1% - поэтому важно поддерживать ее за счет активного транспорта.
К другим электровозбуждаемым каналам относятся Са++ каналы. Они обеспечивают ПД в гладкомышечных клетках, в телах, дендритах и синаптических окончаниях многих нервных клеток.

Теория Ходжкина-Хаксли
Согласно теории Ходжкина-Хаксли, возбуждение элемента мембраны связано с изменениями проводимости мембраны для ионов Na+ и K+. Проводимости мембраны сложным образом зависят от мембранного потенциала и времени
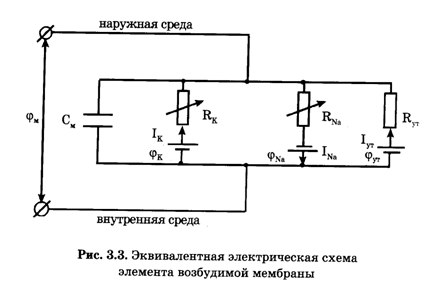
На эквивалентной электрической схеме элемента мембраны равновесные потенциалы Нернста моделируются источниками напряжений с электродвижущими силами jNa, jK, jутечки, а проводимости элемента мембраны для различных ионов - резисторами.
Ходжкин и Хаксли предложили математическую модель, которая описывала изменения проводимостей, а следовательно, и токов ионов Na+ , K+ через мембрану в процессе возбуждения.
Основными постулатами этой модели являются:
1) в мембране существуют отдельные каналы для переноса ионов Na+ , K+
2) во внутренней структуре мембраны существуют некоторые заряженные частицы, управляющие проводимостью каналов. В зависимости от величины напряженности приложенного электрического поля эти гипотетические частицы могут передвигаться в мембране, и тем самым увеличивать или уменьшать потоки ионов Na+ и К+ через каналы (сушествование «воротных токов» было доказано позже)
Предполагается, что ионы калия могут проходить через канал, если к его участку под действием электрического поля подойдут одновременно четыре однозарядные частицы. Обозначим n - вероятность подхода одной такой частицы. Тогда проводимость ионов калия:
gK=gк-n4, (1)
где gк - максимальная проводимость канала для ионов К+. Четвертая степень при n определялась эмпирически. Величина n4 объяснялась как вероятность нахождения одновременно четырех активирующих частиц в некотором определенном участке мембраны.
Изменение проводимости для ионов Na+ описывалось более сложным выражением. Для натриевого канала предполагалось, что он открывается, если одновременно в данный участок попадают три активирующие частицы и удаляется одна блокирующая. Тогда, обозначив m - вероятность прихода активирующей частицы, h- вероятность удаления блокирующей, Получаем:
gNa=gNa-m3h, (2)
где gNa ~ максимальная проводимость канала для ионов Na
Степени при m и h также подбирались эмпирически, чтобы наилучшим образом описать кинетику токов. Численные значения n, m и h имеют смысл вероятности нахождения соответствующей частицы в данном месте канала, а величины их могут меняться от 0 (отсутствие частицы) до 1 (нахождение ее в заданном месте).
Кинетика перераспределения частиц через мембрану при действии электрического поля описывается системой однотипных уравнений (Рубин А.Б. 2том с.176).
Так, для калиевого канала уравнение кинетики записывается в виде:

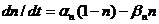 (3)
(3)
где  и
и  - константы скорости перескока частицы, активирующей калиевый канал, к нему и от него соответственно. Константы скорости являются функциями мембранного потенциала
- константы скорости перескока частицы, активирующей калиевый канал, к нему и от него соответственно. Константы скорости являются функциями мембранного потенциала
Уравнение для мембранного тока можно представить в виде (Антонов В.Ф Физика и биофизика с.250):
 (4)
(4)
Для расчета формы потенциала действия решаются уравнения 4 и три уравнения типа 3 для параметров n,m,h. Решение соответствует эксперименту в пределах 10%.
Дата добавления: 2018-04-15; просмотров: 330; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!