Основные направления эволюции водорослей
Все отделы водорослей, кроме красных, ведут свое происхождение
Рис.1. Внешний вид сифональной
водоросли каулерпы
дифференциации,
от различно окрашенных жгутиковых – одноклеточных, активно подвижных в воде организмов, снабженных жгутиками. Одно из самых
т.е. приспособления отдельных участков тела к выполнению своих функций.Многоклеточные водоросли могут иметь нитчатую,
разнонитчатую, пластинчатую формы. Однако их вегетативное тело еще не разделено на ткани. Водоросли живут в относительно стабильных условиях - элементы, необходимые для их питания и развития (вода, минеральные вещества, углекислый газ, кислород), находятся непосредственно в воде и окружают их со всех сторон. Самый сложный уровень организации характерен для бурых водорослей, имеющих клетки, сходные с ситовидными элементами высших растений, однако они еще слабо дифференцированы на ткани. Тип организации вегетативного тела, характерный для всех водорослей называется талломом.
Систематика водорослей
Известно около 40 тыс. видов водорослей. В систематическом отношении они представляют собой совокупность многих отделов, различающихся по набору пигментов и деталям структуры хроматофоров, по продуктам фотосинтеза, накапливающимся в клетке, по строению жгутикового аппарата.
Различают следующие отделы водорослей: О. Красные
|
|
|
О. Бурые
O. Зеленые
О. Золотистые
О. Желто-зеленые О. Диатомовые
О. Эвгленовые
О. Динофитовые и др.
Все отделы в процессе эволюции развивались в основном независимо друг от друга. Причем в большинстве из них встречаются разнообразные типы организации таллома (монадный, амебоидный, коккоидный, сифональный, нитчатый, слоевищный и другие типы), что свидетельствует о параллелизме в эволюции отделов. От водорослей ведут свое происхождение и высшие растения.
OТДЕЛ ЗЕЛЕНЫЕ ВОДОРОСЛИ
Этот отдел включает около 7000 видов, представленных, в основном, пресноводными одноклеточными, нитчатыми, колониальными, слоевищными формами. Зеленые водоросли большей частью обитают в пресноводных водоемах и имеют зеленую окраску из-за преобладания хлорофилла (хлорофиллы а, b) над другими пигментами (каротиноидами). Основное запасающее вещество клеток этих водорослей – крахмал, а в состав клеточной стенки входят целлюлоза, гемицеллюлозы и пектиновые вещества.
Представителями данной группы водорослей являются хламидомонада, спирогира, улотрикс, хлорелла.
Хламидомонада – это одноклеточная подвижная водоросль, живущая в стоячей воде. Ее тело имеет овальную, грушевидную или шаровидную форму. Клетка одета прочной оболочкой. На переднем конце
|
|
|
хламидомонада несет два одинаковых жгутика, с помощью которых она
 активно передвигается в воде. Протопласт содержит одно ядро, обычно чашевидный хроматофор, две пульсирующие вакуоли, красный глазок (стигму). Стигма воспринимает изменения в интенсивности освещения, и водоросль перемещается туда, где интенсивность света оптимальна для фотосинтеза.
активно передвигается в воде. Протопласт содержит одно ядро, обычно чашевидный хроматофор, две пульсирующие вакуоли, красный глазок (стигму). Стигма воспринимает изменения в интенсивности освещения, и водоросль перемещается туда, где интенсивность света оптимальна для фотосинтеза.
восстанавливается гаплоидное состояние, характерное для взрослых организмов. Молодые клетки высвобождаются и дают начало взрослым особям (рис.2, 3-5).
Улотрикс – многоклеточная нитчатая зеленая водоросль, обитающая в проточных водоемах. Ведет прикрепленный образ жизни. Нити водоросли состоят из ряда коротких клеток. В цитоплазме каждой из них расположены ядро и хроматофор в виде незамкнутого кольца.
Взрослое растение улотрикса гаплоидно. Бесполое размножение осуществляется с помощью зооспор. При этом каждая клетка, кроме той, с помощью которой нить прикрепляется, может разделиться на 2 или 4 подвижные зооспоры. Они выходят в воду, плавают, затем прикрепляются к какому-либо подводному предмету и делятся, образуя новые нити улотрикса (рис.3, 1).
|
|
|
Рис.2. Схема жизненного цикла хламидомонады: 1 – образование зооспор; 2 –
образование гамет; 3 – слияние гамет; 4 – зигота; 5 – «прорастание» зиготы
Взрослая особь хламидомонады гаплоидна. Имеет два типа размножения: бесполое и половое. Бесполое размножение осуществляется с помощью зооспор. Материнская клетка сбрасывает жгутики и делится на 2-8 частей путем митоза. Из каждой зооспоры вырастает взрослая хламидомонада (рис.2, 1).
Половой процесс (изогамия, гетерогамия) заключается в образовании и слиянии гамет. Гаметы образуются внутри материнской клетки так же, как зооспоры, но в большем количестве и, соответственно,
меньших размеров (рис.2, 2). Процесс оплодотворения происходит в воде.

Рис.3. Строение клеток зеленых нитчатых водорослей: 1 – улотрикс; 2 - спирогира
В неблагоприятный период в некоторых клетках водоросли образуются многочисленные мелкие одинаковые гаметы со жгутиками. Гаметы выходят в воду и сливаются попарно. Причем обычно сливаются гаметы, возникшие на разных особях. Зигота покрывается толстой оболочкой и может долго находиться в состоянии покоя. При наступлении благо- приятных условий зигота делится мейозом на 4 клетки. Каждая из них способна дать начало новому организму.
|
|
|
Образующаяся зигота покрывается плотной оболочкой и оседает на дно водоема. После периода покоя зигота делится мейотически, при этом
Спирогира - нитчатая неветвящаяся водоросль, живущая в водоемах
с непроточной, но чистой водой. Большинство видов спирогиры –
плавающие формы со слизистыми скользкими нитями. Клетки соединены
торец в торец и идентичны друг другу. В центре клетки расположена большая вакуоль. Хроматофоры – спиралевидные (рис.3, 2). Вегетативное размножение осуществляется частями таллома, то есть путем разрыва нитей на отдельные участки или даже отдельные клетки.
Половой процесс – конъюгация. При половом размножении две нити располагаются параллельно друг другу и супротивные клетки обеих нитей соединяются при помощи копуляционных выростов или мостиков. Оболочки их в месте соприкосновения растворяются, и образуется сквозной канал, через который сжавшееся содержимое клетки одной нити перетекает в клетку другой и сливается с ее протопластом. Образующаяся в результате оплодотворения зигота после периода покоя прорастает. Этому предшествует редукционное деление ядра: из четырех получившихся ядер три отмирают, а одно остается ядром единственного проростка, выходящего через разрыв наружных слоев оболочки зиготы.
Хлорелла – шаровидная одноклеточная водоросль, встречающаяся в пресных водоемах, почве. Клетки одеты оболочкой, содержат чашевидный хроматофор. Половой процесс отсутствует. При бесполом размножении материнская клетка делится на 4-16 клеток. Образуются неподвижные споры, которые входят наружу после разрыва материнской оболочки.
Хлорелла характеризуется высокими темпами размножения. Каждая клетка в течение суток может образовывать до 10 дочерних клеток. Богата белками, неприхотлива. Широко используется в научных исследованиях.
OТДЕЛ КРАСНЫЕ ВОДОРОСЛИ
Красные водоросли (багрянки) представляют собой древнюю группу многоклеточных организмов. В некоторых современных системах их выделяют в отдельное подцарство. Вегетативное тело у багрянок в большинстве случаев имеет вид разветвленных слоевищ красного цвета,
прикрепленных к субстрату с помощью ризоидов.
В хроматофорах красных водорослей содержатся такие пигменты, как хлорофиллы a, d, синий фикоцианин, красный фикоэритрин, которые и обусловливают их ярко-малиновую окраску. В клеточных оболочках часто откладывается карбонат кальция. Основное запасающее вещество – багрянковый крахмал – по составу более близок к гликогену (запасному веществу животных), нежели к крахмалу.
При бесполом размножении у красных водорослей вместо зооспор из содержимого спорангия образуются голые, лишенные подвижности клетки. Если они возникают по одной в каждом спорангии, их называют моноспорами, у большинства же представителей образуется по 4 клетки - тетраспоры. Споры, попав в воду, прикрепляются к субстрату и прорастают в новые особи.
Половой процесс у багрянок – оогамный. При этом на слоевище образуются половые органы – одноклеточные антеридии и оогонии. Мужские гаметы лишены жгутиков и пассивно переносятся к женскому оогонию, внутри которого происходит их слияние с яйцеклеткой.
Красные водоросли являются обитателями теплых тропических морей и живут на большой глубине. Например, кораллиновая водоросль однажды была найдена на глубине 268 м. Багрянка порфира активно используется в пищу и специально выращивается на морских плантациях Японии.
OТДЕЛ БУРЫЕ ВОДОРОСЛИ
Бурые водоросли – многоклеточные организмы, обитающие в морях. Это самые крупные из известных водорослей. Представлены, в основном, нитчатыми и слоевищными формами.
Пигментами бурых водорослей являются хлорофиллы а, с, каротиноиды и фукоксантин, избыток которого придает бурую окраску этим водорослям. Запасающие вещества представлены полисахаридами
ламинарином, маннитом и жирами. Половой процесс – оогамный с чередованием гаплоидной и диплоидной ядерных фаз.

Рис.4. Представители бурых водорослей: 1 – ламинария; 2 –
саргассум; 3 - макроцистис
Известным представителем этой группы является ламинария (морская капуста), произрастающая в наших дальневосточных и северных морях. Ламинария состоит из листоподобных пластинок, ножки и прикрепляется к субстрату с помощью ризоидов (рис.4, 1). Часто образует заросли. Особи достигают от 5 до 50 м в длину.
Взрослое растение ламинарии является спорофитом (диплоидное поколение). На слоевище возникают зооспорангии, в которых в результате мейоза образуются зооспоры. Зооспоры попадают в воду и прорастают в заростки (гаметофиты). На одних гаплоидных заростках образуются женские половые органы – оогонии с яйцеклетками, на других – мужские антеридии со сперматозоидами. После слияния
сперматозоида с яйцеклеткой образуется диплоидная зигота, которая сразу же прорастает во взрослую особь – спорофит. Жизненный цикл ламинарии сходен с высшими растениями.
Ламинария богата питательными веществами, в том числе йодом, и используется человеком в пищу. Ею также питаются многочисленные морские обитатели – моллюски, рыбы, морские ежи. Последние способны уничтожить большие заросли.
Чаще бурые водоросли растут на глубине 6-15 м и ниже. Наиболее знаменито скопление бурых водорослей саргассум посреди Атлантического океана. На плаву саргассумы удерживаются благодаря особым воздушным пузырькам (рис.4, 2). Эти плавающие водоросли часто сгоняются ветром в кучи, сцепляются между собой и образуют длинный «саргассовый эшелон», которой может достигать в длину 5-7 км. Раньше некоторые моряки считали, что в зарослях саргассума можно увязнуть. К числу бурых водорослей относится и «Голиаф растительного мира» - макроцистис, который может достигать 60 м в длину, а каждые сутки вырастать на полметра (рис.4, 3).
Значение водорослей
Повсеместное распространение водорослей определяет их большое значение в природе и хозяйственной деятельности человека.
Водоросли создают огромное количество биомассы. За счет органического вещества, образуемого водорослями, живет животный мир озер, рек, морей и океанов. В процессе фотосинтеза водоросли освобождают громадное количество кислорода, который не только растворяется в воде, но и выделяется в атмосферу.
Водоросли являются источниками для получения многих полезных продуктов:
Y альгиновой кислоты (бурые водоросли), которая идет на изготовление гелей, кремов, эмульгаторов для мороженого, красок, лаков
и т.д. ;
Y полисахарида агар-агара (красные водоросли), используемого в микробиологии для приготовления плотных питательных сред, в пищевой промышленности - для приготовления мармелада, пастилы;
Y удобрений (красные и бурые водоросли).
Такие богатые йодом водоросли, как ламинария и порфира, являются ценными пищевыми продуктами и используются как в сыром виде, так и для приготовления различных блюд.
Микроскопические водоросли иногда применяются для очистки сточных вод. Некоторые водоросли (например, хлорелла) способны накапливать радионуклеиды, что может использоваться для дополнительной очистки слабоактивных сточных вод атомных электростанций.
Одноклеточным водорослям присущи все характерные признаки типичных растений, поэтому они являются идеальным материалом для научных исследований, так как обладают высокой скоростью размножения. Примером таких водорослей может служить хлорелла, которой по праву принадлежит почетное место в исследованиях фотосинтеза.
Сильное размножение водорослей может наносить ущерб хозяйственной деятельности человека. Вместе с другими организмами они обрастают морские суда, ухудшая их эксплуатационные качества. Некоторые водоросли, особенно сине-зеленые, вызывают «цветение» воды. «Цветение» наблюдается при достаточно теплой погоде, когда в воде много питательных веществ. Такая ситуация часто искусственно создается человеком, когда в воду сбрасывают промышленные отходы или же когда в реки и озера попадают удобрения с полей. В результате начинается взрывоподобное размножение водорослей и они, в нарушение всех законов природы, начинают отмирать раньше, чем их успевают съесть. При последующем разложении органических остатков происходит
столь же интенсивное размножение аэробных бактерий и вода почти
полностью лишается кислорода. Все это происходит очень быстро, и из-за нехватки кислорода начинают гибнуть рыбы и другие животные и растения. Гибели животных также способствуют токсины, выделяемые при «цветении» сине-зелеными и другими водорослями. Увеличение концентрации питательных веществ в воде, которое запускает весь этот процесс, называется эвтрофизацией водоема.
Вопросы для самоконтроля
1. Какими признаками характеризуются низшие растения?
2. Почему оогамный половой процесс является наиболее эволюционно продвинутым?
3. Почему водоросли не имеют тканевого строения?
4. Какие признаки являются основополагающими при классификации водорослей?
5. Какое поколение является доминирующим в жизненном цикле хламидомонады?
6. Какими признаками характеризуются бурые водоросли?
7. Каковы причины «цветения» водоемов?
ПОДЦАРСТВО ВЫСШИХ РАСТЕНИЙ
Высшие растения, как и все живые существа, имели водных предков, и их эволюция неразрывно связана с завоеванием суши. Большинство ученых считает, что предками наземных растений были зеленые водоросли. С этой группой водорослей высшие растения сближает сходство пигментного состава (хлорофиллы а, в и каротиноиды), строение репродуктивных органов. Основное запасающее вещество у этих двух групп организмов - крахмал, который откладывается в пластидах, а главным веществом клеточной стенки является целлюлоза. Некоторые ученые склоняются к мнению, что высшие растения произошли от бурых или красных водорослей, у которых так же, как и у наземных растений, имеется оогамный половой процесс и сложно устроенные гаметангии. Кроме того, жизненный цикл некоторых бурых водорослей сходен с циклом воспроизведения наземных растений. Предполагается, что толчком к выходу на сушу растений послужило достаточное накопление в атмосфере кислорода, появление озонового слоя, а также усиление конкуренции в морях между организмами за источники питания и свободное место.
Выход на сушу стал поворотным моментом в эволюции всех растительных организмов, которым пришлось осваивать совершенно новую среду обитания, совершенно новые условия. Естественно, что приспособление к этим новым условиям повлекло за собой и резкие изменения в строении тела, размножении и образе жизни растений.
С какими же трудностями сталкивается растение при переходе к наземному образу жизни? Во-первых - это проблема обезвоживания. Представьте, что произойдет с водорослью, если ее вынести из воды. Поэтому у высших растений появляются покровные ткани и кутикула, препятствующие излишнему испарению воды и защищающие от механических воздействий. Если в водной среде растение всасывало воду
всей поверхностью, то на суше появилась необходимость образования
корнеподобных структур для извлечения воды из почвы и прикрепления к субстрату. Полученную из влажной почвы воду и растворенные в ней минеральные вещества нужно поднять на высоту растения, поэтому появляются проводящие ткани. Так как в воздушной среде резко возрастают механические нагрузки, то возникает потребность образования механических тканей для поддержания тела растения. Необходимость газообмена, которая в наземных условиях происходит с воздушной средой, а не с раствором, привела к образованию устьиц, расположенных в эпидермисе. Для обеспечения процесса фотосинтеза потребовалось образование ассимиляционной ткани. Таким образом, наземные растения выработали различные ткани, а клетки, слагающие их, подверглись сильной дифференциации, в результате чего стали выполнять более узкие функции лучше, чем недифференцированные клетки водорослей.
Настоящие вегетативные органы впервые возникают только у семенных растений, поэтому структуры, выполняющие аналогичные функции у споровых растений, называть корнями, стеблями и листьями, на наш взгляд, неправомерно. В настоящем пособии мы приводим традиционные названия вегетативных органов споровых растений, однако указываем их в кавычках.
Органы размножения у высших растений также подверглись изменению и усложнению. Половые органы, производящие гаметы, являются многоклеточными структурами и имеют защитную оболочку. Антеридии (мужские гаметангии) обычно представляют собой замкнутое тело, окруженное оболочкой, внутри которого находится многоклеточная сперматогенная ткань, дающая начало сперматозоидам или безжгутиковым спермиям. Архегонии (женские гаметангии) часто имеют колбовидную форму и состоят из брюшка с яйцеклеткой внутри и шейки с канальцевыми клетками. В процессе эволюции от риниофитов до
 цветковых растений антеридии и архегонии постепенно подвергались редукции и полному исчезновению.
цветковых растений антеридии и архегонии постепенно подвергались редукции и полному исчезновению.
В наземных условиях у высших растений, еще не перешедших к семенному размножению, процесс полового размножения всегда связан с водой (по ней сперматозоиды передвигаются к яйцеклетке), и его осуществление всегда находится под угрозой. Высшие растения, вышедшие на сушу, еще долго сохраняют древнюю связь с водой, унаследованную от своих водорослевых предков.
При бесполом размножении высших растений в многоклеточных спорангиях образуются споры, распространяющиеся в воздушной среде и покрытые плотной оболочкой, защищающей их от высыхания.
Для всех высших растений характерна правильная смена поколений: гаплоидного гаметофита и диплоидного спорофита. На гаметофите в результате митотического деления образуются гаметы – сперматозоиды или яйцеклетки. Сливаясь, гаметы формируют диплоидную зиготу, которая дает начало многоклеточному зародышу, развивающемуся в следующее поколение - спорофит. На спорофите, в результате редукционного деления, образуются гаплоидные споры, т.е. здесь мы видим возврат к гаплоидному поколению. Пример жизненного цикла приведен на рис. 5.
Такая смена поколений появилась уже у ряда водорослей (бурых и некоторых зеленых) и от водорослей же была унаследована высшими растениями. У подавляющего большинства высших растений в жизненном цикле доминирует спорофит, подобно тому, как это наблюдается у ряда бурых водорослей. Однако другой путь эволюции высших растений привел к формированию моховидных, у которых преобладающим поколением является гаметофит, что свойственно некоторым зеленым водорослям.


Рис.5. Обобщенная схема жизненного цикла равноспорового растения
Гаметофиты наземных растений, как правило, представляют собой небольшие растения, не дифференцированные на ткани и органы, чем очень напоминают водоросли. Они весьма чувствительны к недостатку влаги, могут жить в условиях сильного увлажнения и легко гибнут при пересыхании. Обычно гаметофиты недолговечны и только у самых древних высших растений развиваются в течение нескольких лет. У всех разноспоровых растений они созревают очень быстро, иногда за несколько часов, и крайне редуцированы – представлены лишь небольшим количеством клеток. Следовательно, эволюция жизненного цикла высших растений направлена на сокращение стадии гаметофита как более уязвимого поколения.
Высшие растения в настоящее время господствуют на Земле, составляя основную массу зеленого покрова. Они насчитывают не менее 300000 видов и распространены на всех континентах, во всех
климатических зонах, как в широтном направлении, так и от уровня моря до границы вечных снегов.
Подцарство высших растений делится на 9 отделов. Здесь мы рассмотрим лишь основные из них. Отдел моховидные относится к бессосудистым споровым растениям (т.е. у мхов отсутствует хорошо развитая проводящая система, а размножение бесполым путем осуществляется с помощью спор). Отделы риниофиты, плауновидные, хвощевидные и папортниковидные являются споровыми сосудистыми растениями, а голосеменные и цветковые - семенными сосудистыми растениями.
ОТДЕЛ РИНИОФИТЫ (ПСИЛОФИТЫ)
 Предполагается, что первые растения, вышедшие на сушу, имели небольшие размеры, простое строение и произрастали в прибрежной полосе. Самыми примитивными и древними из известных науке высших растений является вымерший отдел риниофиты.
Предполагается, что первые растения, вышедшие на сушу, имели небольшие размеры, простое строение и произрастали в прибрежной полосе. Самыми примитивными и древними из известных науке высших растений является вымерший отдел риниофиты.
Риниофиты появились в позднем силуре (420 млн лет назад) и вымерли в
среднем девоне (около 380 млн лет назад).
группа, от которой произошли моховидные, плауновидные, хвощевидные и папоротниковидные растения.
ОТДЕЛ МОХОВИДНЫЕ
Моховидные, или мхи, - наиболее примитивные из высших растений, известны с карбона. Всего насчитывается около 25000 видов. Поселяются повсюду, кроме морей, засоленных почв и местообитаний, подверженных сильной эрозии. На моховых болотах образуют основную массу торфяных залежей.
Мхи - обычно многолетние, небольшие (от нескольких мм до нескольких см), реже крупные (до 60 см и более) растения. Тело представляет собой слоевище (антоцеротовые, некоторые печеночники) или расчленено на “стебель” и “листья” (“листостебельные”). Корней у мохообразных нет, всасывание воды и прикрепление к субстрату осуществляется выростами клеток - ризоидами, проводящая система, как правило, отсутствует. Наиболее характерным признаком мохообразных, отличающим их от остальных высших растений, является жизненный цикл, в котором преобладает гаметофит (половое поколение). У мхов сильно развита способность к вегетативному размножению, что является одной из причин образования групповых форм роста типа ковриков и подушек.
Жизненный цикл моховидных происходит по уже известной вам
Рис. 6. Строение спорофита ринии большой
Отдел включает споровые растения,
схеме (рис.5). На хорошо развитом фотосинтезирующем гаметофите
состоящие из простых, дихотомически ветвящихся осей, рост которых осуществлялся с помощью верхушечных меристем. Дифференциация на стебли, корни, листья отсутствовала (рис.6). На верхушках осей у многих представителей располагались одиночные крупные спорангии.
Название отдела происходит от деревни Райни (Шотландия), близ которой остатки одного из этих древних растений прекрасно сохранились в каменистых породах. Считают, что риниофиты - исходная предковая
развиваются половые органы - архегонии и антеридии. В антеридиях образуется большое число двужгутиковых сперматозоидов. В каждом из архегониев созревает по одной яйцеклетке. В сырую погоду (во время дождя) сперматозоиды, передвигаясь в воде, проникают к яйцеклетке, находящейся внутри архегония. Один из них сливается с ней. Из оплодотворенной яйцеклетки (зиготы) образуется спорофит, т.е. бесполое поколение, представленное коробочкой, сидящей на ножке. В коробочке
созревают споры, причем при их образовании происходит мейоз, т.е.
каждая спора имеет гаплоидный набор хромосом. При попадании споры в благоприятные условия у некоторых мхов образуется ветвящаяся нить или пластинка – протонема, на которой формируется гаметофит.
Отсутствие проводящей системы и преобладание в жизненном цикле гаметофита резко отличает моховидные от других высших растений. У ботаников не существует единого взгляда на происхождение этих растений. Одни считают, что мхи произошли от зеленых водорослей независимо от остальных высших растений, а другие - от риниофитов в результате редукции проводящих тканей и перестройки жизненного цикла.
В отделе моховидных выделяют 3 класса: антоцеротовые, печеночники и листостебельные мхи. Антоцеротовые мхи часто относят к самостоятельному отделу, так как у этой группы растений имеются признаки, роднящие их с водорослями и не свойственные высшим растениям. Мы подробнее рассмотрим класс листостебельных мхов на примере двух представителей - сфагнума и кукушкина льна.
Сфагновые мхи.Существует примерно 350 видов мхов, относящихся к единственному роду сфагнум. Сфагновые мхи широко распространены в умеренных и холодных областях Северного полушария, образуя сплошной покров на болотах и во влажных лесах. Торфяные болота занимают 1% территории суши.
Гаметофиты сфагнума образуют крупные светло-зеленые или красноватые куртинки. “Стебли” прямостоячие, с пучковидно расположенными олиственными “ветвями”. На верхушке “ветки” укорочены и собраны в довольно плотную головку (рис.7, А). Наружный слой “стеблей” состоит из крупных бесцветных водоносных клеток с порами, лишенных живого содержимого. “Листья” сфагнума однослойные, имеют клетки двух типов: фотосинтезирующие и водоносные. Фотосинтезирующие клетки червеобразной формы, в них
содержатся хлоропласты. Они расположены между широкими
водоносными клетками. Благодаря множеству водоносных клеток сфагнум может всасывать большое количество воды (в 20 раз больше собственной сухой массы).
Спорофит сфагнума представлен округлой формы коробочкой, в которой образуются споры, и крышечкой (рис.7, В, С). При созревании спор в результате увеличения давления внутри коробочки, крышечка отскакивает и споры вылетают наружу. Это происходит обычно в теплую солнечную погоду, что обеспечивает лучшее распространение спор.
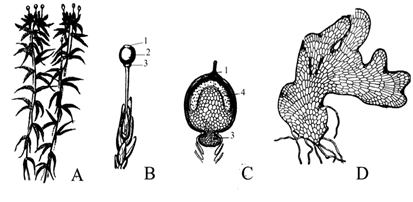
Рис.7. Строение сфагнума: А – гаметофит с развившимися спорофитами; B – внешний вид спорофита; C – внутреннее строение спорофита; D - протонема: 1 – крышечка, 2 – коробочка, 3 – ножка, 4 – спорангий
Растут сфагновые мхи очень медленно - всего по 2-3 см в год. Эти растения содержат бактерицидное вещество - сфагноль, которое подавляет деятельность почвенных микроорганизмов и способствует накоплению неразложившихся органических остатков (торфа).
Благодаря своим бактерицидным и абсорбирующим свойствам сфагновые мхи использовались в Европе с 1880-х гг. как перевязочный материал для ран и нарывов. В настоящее время торф используют в качестве топлива и удобрения полей. Также торф находит применение в
химической промышленности для изготовления воска, парафина, фенола, карболовой кислоты.
Ярким представителем зеленых мхов является кукушкин лен. “Стебель” кукушкина льна покрыт жесткими, темно-зелеными шиловидными “листьями”, имеет ризоиды и достигает высоты 30-40 см. В “стебле” имеется проводящая система довольно примитивного строения. Гаметофиты этого растения раздельнополы (рис.8, А). На верхушке
«стеблей» образуются антеридии и архегонии. После оплодотворения из зиготы развивается спорофит, представленный коробочкой на длиной ножке, в которой созревают гаплоидные споры. Сверху коробочка прикрыта легко опадающим колпачком с тонкими, направленными вниз
Рис.8. Строение кукушкина льна: А – женский гаметофит со спорофитами; Б – продольный разрез коробочки; В – поперечный разрез коробочки; 1 – урночка, 2 – крышечка, 3 – кольцо, 4 – спорангий, 5 – колонка, 6 – шейка
крышечки образуется кольцо, состоящее из клеток с неравномерно утолщенными стенками (рис.8, Б, В). По этому кольцу происходит отделение крышечки от урночки (ее
сбрасывание).
волосками, напоминающими льняную пряжу, отсюда и название растения.
 Коробочка имеет сложное строение и подразделяется на крышечку, урночку и шейку. Внутри
Коробочка имеет сложное строение и подразделяется на крышечку, урночку и шейку. Внутри
коробочки имеется колонка, состоящая из бесплодных клеток.
Вокруг колонки расположен спорангий. На границе урночки и
Распространение спор происходит в сухую погоду. Спора попадает в почву и прорастает в протонему - ветвящуюся нить, на которой формируется гаметофит. Встречается кукушкин лен, большей частью, в смешанных и еловых лесах.
Мхи играют важную роль на болотах, в тундровых, лесных сообществах, создавая особый водный режим почвы и пополняя запасы органическим веществом.
Вопросы для самоконтроля
1. Каковы причины выхода растений на сушу?
2. Назовите причины появления у растений тканевого строения.
3. Почему у большинства высших растений доминирующим поколением является спорофит?
4. Какими примитивными признаками обладали представители отдела риниофитов?
5. Назовите характерные признаки представителей отдела моховидных.
6. В чем отличие жизненного цикла мхов от других высших
растений?
7. Чем представлен спорофит у моховидных?
8. Какое строение имеет коробочка кукушкина льна?
ОТДЕЛ ПЛАУНОВИДНЫЕ
Плауновидные - одна из наиболее древних групп высших растений. Появились в начале девона палеозойской эры. Их предками, вероятно, были риниофиты.
Современные плауны представлены многолетними травянистыми вечнозелеными растениями. Доминирующим поколением является спорофит, который представлен стелющимися и приподнимающимися осями, имеющими дихотомическое ветвление (рис.9). Нарастание осей происходит вследствие деятельности верхушечных меристем. Но, в отличие от риниоподобных предков, у плаунов появляются выросты поверхностных тканей - микрофиллы, которые подразделяются на трофофиллы - фотосинтезирующие микролистья - и спорофиллы, защищающие спорангии от внешних воздействий. Трофофиллы значительно увеличивают фотосинтезирующую поверхность.

Рис.9. Плаун булавовидный: спорофит с колоском
У большинства представителей плауновидных на верхушке осей располагаются спорангии, собранные в колоски (стробилы). Спороносный колосок состоит из оси и спорофиллов, на которых располагаются округлые спорангии. Образование спор происходит в результате редукционного деления (мейоза). Среди плауновидных встречаются равноспоровые и разноспоровые растения. Этот признак очень важен при делении отдела на два класса: плауновых и полушниковых.
У представителей класса плауновые в спорангиях созревают морфологически одинаковые споры, т.е. они относятся к равноспоровым растениям.
Из споры, попавшей в благоприятные условия, образуется заросток (гаметофит). Заросток плауна многолетний, имеющий вид маленького клубенька (2-3 мм) с ризоидами. Он бесцветный, лишен хлорофилла и самостоятельно питаться не может. Его рост начинается только после проникновения в тело гриба. Заросток развивается, как правило, в почве в
течение 10-15 лет. На верхней стороне гаметофита образуются антеридии и архегонии. Оплодотворение происходит при наличии воды. Из оплодотворенной яйцеклетки развивается зародыш, вырастающий в многолетнее вечнозеленое растение - спорофит. Наиболее типичными представителями этого класса являются плаун булавовидный (рис.9) и плаун годовалый, произрастающие в хвойных лесах.
Род селягинелла, относящийся к классу полушниковых, характеризуется разноспоровостью. В одном и том же колоске селягинеллы имеются спорангии двух типов. Одни из них более крупные (мегаспорангии) и содержат по 4 крупных мегаспоры. Другие спорангии мельче (микроспорангии), в них содержатся многочисленные микроспоры. Содержимое микроспоры превращается в мужской заросток, состоящий из одной вегетативной клетки и одиночного антеридия. Заросток не способен к фотосинтезу и целиком зависит от запаса питательных веществ в микроспоре. Из мегаспоры образуется женский гаметофит с многочисленными архегониями, который питается в основном за счет запасов споры. Передвижение сперматозоидов происходит в воде после дождя или росы. Из оплодотворенной яйцеклетки со временем вырастает новое растение - спорофит. Схема жизненного цикла селягинеллы показана на рис. 10.
Характерной особенностью разноспоровых растений является значительная редукция гаметофита. Зрелые гаметофиты селягинеллы не отделены от спорофита и этим отличаются от всех уже ранее рассмотренных нами сухопутных растений. Это очень важное эволюционное достижение - одно из приспособлений к жизни на суше, которое привело к тому, что легко уязвимый гаметофит, не дифференцированный на ткани, стал хотя бы отчасти защищен спорой. Гаметофитное поколение не способно жить самостоятельно, оно питается за счет веществ, которое запасло в споре предшествующее спорофитное
поколение. Как вы увидите дальше, эволюция высших растений направлена на сокращение стадии жизни гаметофита.
Практическое значение плаунов невелико. Некоторые виды плауна содержат парализующий яд, сходный по действию с кураре. Очевидно, поэтому позвоночные животные не употребляют плауны в пищу. Их споры используют в качестве порошка для присыпок, а также для обсыпки пилюль. Плауны способны извлекать из лесной подстилки и торфянистой почвы труднодоступные для других растений питательные вещества, ускоряя круговорот веществ в лесу.

Рис.10. Схема жизненного цикла разноспорового растения
ОТДЕЛ ХВОЩЕВИДНЫЕ
В геологическом прошлом представители этого отдела были весьма разнообразны. Ископаемые хвощевидные, например каламиты, наряду с
древовидными плауновидными, образовывали леса каменноугольного периода. Современные хвощи представлены в растительном мире единственным родом - хвощ.
Строение хвощевидных и их жизненный цикл можно рассмотреть на примере хвоща полевого (рис.11). Это многолетнее травянистое растение, встречающееся в условиях избыточного увлажнения. На “стебле” хвоща вздутые узлы чередуются с ребристыми междоузлиями, отчего “стебель” кажется членистым. К узлам прикрепляются мутовки боковых осей второго порядка. Микрофиллы также расположены мутовчато, причем они срастаются между собой почти до самой верхушки, охватывая основание междоузлия “стебля”, и выполняют функцию защиты вставочных меристем. Микрофиллы хвощей утратили функцию фотосинтеза. Как мы видим, у хвощей, в отличие от плаунов и риниофитов, кроме верхушечных, появляются еще и вставочные меристемы, за счет которых растение способно расти дальше при повреждении верхушки.
Снаружи “стебель” хвоща покрыт эпидермисом, клеточные стенки которого пропитаны кремнеземом, придающим “стеблям” известную жесткость и препятствующим поеданию животными. Под эпидермисом располагается фотосинтезирующая ткань, проводящие пучки, в центре находится большая воздухоносная полость. Подземная часть представлена стелющимися осями (“корневищами”), которые несут в узлах придаточные корни. С помощью “корневищ” осуществляется вегетативное размножение растения.

Рис.11. Хвощ полевой: 1 – вегетативный «побег»; 2 – спороносный побег; 3 – спорангиофор; 4 - споры с элатерами; 5 – мужской гаметофит; 6
– женский гаметофит
Весной из глубоко залегающих “корневищ” хвоща полевого вырастают оси, заканчивающиеся спороносными колосками (рис.11, 2). Каждый колосок состоит из оси, перпендикулярно которой крепятся шестиугольные щитки – спорангиофоры (рис.11, 3), под которыми находятся 6-10 спорангиев, содержащих гаплоидные споры. Между щитками образуются щели, через которые высыпаются созревшие споры. Каждая спора имеет два придатка в виде узких лент - элатер, способствующих разрыхлению и распространению спор (рис.11, 4).
Морфологически споры одинаковы, но отличаются физиологически. Одни из них, попав в более благоприятные условия, образуют женские заростки, другие - мужские. Мужской заросток представляет собой
маленькую, размером в несколько миллиметров зеленую пластинку, расчлененную на лопасти и прикрепленную ризоидами к почве (рис.11, 5). На концах лопастей развиваются антеридии, содержащие многожгутиковые сперматозоиды. Женский заросток крупнее, он несет архегонии (рис.11, 6). Оплодотворение происходит при наличии воды. Из зиготы развивается многолетний спорофит. У хвоща полевого после рассеивания спор спороносные “побеги” отмирают, и на смену им вырастают зеленые ассимилирующие оси (рис.11, 1). Некоторые виды хвоща имеют только один тип “побега”. Он одновременно и спороносный и фотосинтезирующий.
Хвощ полевой применяют как кровоостанавливающее и мочегонное средство. Это растение известно садоводам как злостный сорняк.
ОТДЕЛ ПАПОРОТНИКОВИДНЫЕ
Папоротниковидные также являются древними растениями. Значительная часть их вымерла. В настоящее время отдел папоротниковидные включает около 10000 видов. Во флоре нашей страны насчитывается более 100 видов.
Представители этого отдела очень разнообразны по внешнему виду,
вид;
 Рис.12. Щитовник мужской: 1 – общий
Рис.12. Щитовник мужской: 1 – общий
2 – часть листа с сорусами
Папоротники растут, большей частью, в сырых тенистых лесах, по лесным оврагам, кустарникам. В отличие от плауновидных и хвощевидных, для папоротников характерно развитие вай («макро- листьев»), которые могут достигать в высоту до 1 м и более. Вайя состоит из фотосинтезирующей пластики и черешка, который крепится к мясистому “корневищу” (рис.12, 1). “Листовые” пластинки довольно часто перистые, нередко многократно рассеченные.
жизненным формам, условиям обитания. Наибольшее их разнообразие характерно для влажных тропических лесов. Здесь встречаются древовидные папоротники, достигающие в высоту до 25 метров, а в диаметре - 50 см. В умеренных зонах произрастают только травянистые многолетние папоротники. Исключением является сальвиния плавающая - водное однолетнее растение.
Вайи имеют неограниченный рост и растут верхушкой, чем отличаются от настоящего листа семенных растений. Формирование вайи происходит медленно. Молодая вайя вначале улиткообразно свернута и скрыта под поверхностью почвы. По мере роста она разворачивается и на третий год полностью распрямляется. Предполагают, что вайя произошла в процессе эволюции от системы уплощенных и сросшихся боковыми частями осей риниоподобных предков, в отличие от микрофиллов хвощей и плаунов, являющихся выростами поверхностных тканей. Появление вайи существенно увеличило фотосинтезирующую поверхность, что дало значительное преимущество папоротникам в конкурентной борьбе за
“место под солнцем” с хвощами и плаунами.
Отдел папоротниковидные включает 8 классов, 5 из них представлены ископаемыми формами. Современные папоротники разделяют на 3 класса: ужовниковые, мараттиевые и многоножковые.
Почти все папоротники, за исключением водных, являются равноспоровыми растениями. Рассмотрим цикл развития равноспорового папоротника на примере щитовника мужского, являющегося представителем класса многоножковых. Этот папоротник имеет толстое ползучее “корневище”, на конце которого каждый год образуется розетка перисторассеченных вай (рис.12, 1). На нижней поверхности вайи расположены спорангии, собранные в кучки, называемые сорусами (рис.12, 2). Снаружи сорусы порыты покрывальцем, выполняющим функцию защиты. Расположение сорусов на нижней поверхности “листа” более выгодно, чем с верхней, так как обеспечивает лучшую защиту спорангиев, способствует распространению и не мешает фотосинтезу. Внутри спорангия образуются гаплоидные споры. Попав на землю, спора прорастает и образуется заросток (гаметофит). Он представляет собой сердцевидную зеленую пластинку размером около 1 см, прикрепляющуюся к субстрату с помощью ризоидов. На нижней поверхности заростка образуются архегонии и антеридии. В антеридиях развиваются многожгутиковые сперматозоиды. Оплодотворение происходит при наличии воды. Из оплодотворенной яйцеклетки (зиготы) постепенно вырастает многолетний крупный спорофит.
Представляют интерес разноспоровые водные папоротники. Примером может служить сальвиния плавающая, также относящаяся к классу многоножковых (рис.13). Это небольшое плавающее растение, в микро- и мегаспорангиях которого созревают микро- и мегаспоры (рис.13, 2). Из микроспор развиваются сильно редуцированные мужские гаметофиты, состоящие из двух вегетативных клеток и двух антеридиев, в каждом из которых образуется всего по 4 сперматозоида. Женский
гаметофит, находящийся внутри мегаспоры, состоит из нескольких
клеток, заполненных питательными веществами, и 3-5 архегониев. При слиянии сперматозоида и яйцеклетки образуется зигота, дающая начало диплоидному спорофиту. В целом жизненный цикл сальвинии сходен с циклом развития селягинеллы (см. рис.10).

Рис.13. Сальвиния плавающая: 1 – общий вид спорофита; 2 – разрез сорусов; а – микроспорангии, б - мегаспорангии
Значение папоротников достаточно велико. Они являются важнейшим компонентом многих лесных сообществ. Щитовник мужской обладает антигельминтным действием, а молодые, закрученные в улитку вайи орляка обыкновенного используют в пищу.
Таким образом, рассмотренные нами отделы споровых сосудистых растений - риниофиты, плауновидные, хвощевидные и папоротниковидные - обладают следующими общими чертами:
1) наличием проводящей ткани (флоэмы и ксилемы);
2) доминированием в жизненном цикле спорофита;
3) сходным типом жизненного цикла;
4) самостоятельно живущим гаметофитом (за исключением неко- торых разноспоровых растений);
5) бесполым размножением с помощью спор;
6) сходным строением архегониев и антеридиев;
7) необходимостью воды для оплодотворения (по ней подвижные сперматозоиды перемещаются к неподвижной яйцеклетке). Последнее условие значительно ограничивает возможности широкого распространения этих растений.
Вопросы для самоконтроля
1. Почему плауны, хвощи, папоротники называют споровыми сосудистыми растениями?
2. Где развиваются споры у селягинеллы?
3. Какова функция гаметофитов у высших споровых растений?
4. Какие преимущества дает разноспоровость высшим растениям?
5. Какова роль различных представителей споровых растений в природе и хозяйстве?
СЕМЕННЫЕ РАСТЕНИЯ
Одним из важнейших приобретений в эволюции сосудистых растений было семя. Оно явилось, вероятно, одним из факторов, определивших господство семенных растений в современной флоре. Причина этого проста: семя способствует выживанию. Среди высших растений только два отдела характеризуются наличием семени - голосеменные и покрытосеменные. Для этих двух отделов характерны следующие общие признаки:
Y разноспоровость, т.е. имеются два типа спор (микроспоры и мегаспоры);
Y женский гаметофит остается целиком закрытым в
семязачатке (мегаспорангии); семя - это оплодотворенный семязачаток;
Y в жизненном цикле доминирует спорофит, гаметофит крайне редуцирован;
Y для полового размножения не нужна вода, потому что мужские гаметы (спермии) не способны плавать (исключение составляют наиболее примитивные голосеменные); к яйцеклетке спермии передвигаются по пыльцевой трубке;
Y образование побега, состоящего из стебля, листа и почки; почка предохраняет побег, находящийся в зачаточном состоянии, от неблагоприятных воздействий.
По сравнению с высшими споровыми, семенные представляют собой высокий уровень организации растений. Именно у них впервые появляются настоящая корневая система, побег, основным средством расселения видов является семя, а половой процесс не зависит от воды. Благодаря такой независимости семенные растения получили возможность расселиться по всей Земле и стать господствующей группой растений.
ОТДЕЛ ГОЛОСЕМЕННЫЕ
Голосеменные имеют широкое распространение по всему Земному шару. Леса из многих видов голосеменных составляют примерно треть всех лесов планеты. В основном представители этого отдела являются вечнозелеными, реже - листопадными деревьями и кустарниками, иногда - лианами. Листья голосеменных сильно варьируют по форме, размерам, по морфологическим и анатомическим особенностям. Так, по форме листья могут быть чешуевидными, игольчатыми, перистыми и т.д. Для этой группы растений характерно открытое расположение семяпочек (отсюда название отдела). Предполагают, что произошли голосеменные от
вымерших разноспоровых папоротниковидных, объединяемых в группу проголосеменных папоротников.
Отдел голосеменные включает 6 классов, причем классы семенные папоротники и беннеттитовые полностью вымерли. Ныне живущие голосеменные, насчитывающие около 700 видов, относятся к классам саговниковых, гинкговых, гнетовых и хвойных.
В школьном курсе рассматривается только один класс - хвойные, в котором насчитывается около 560 современных видов. К хвойным относятся такие широко известные роды, как сосна, ель, пихта, лиственница, можжевельник, кипарис и менее известные гиганты растительного мира - секвойя, секвойядендрон, метасеквойя. Анатомическое строение стволов хвойных довольно однообразно. Основную часть стебля занимает древесина (вторичная ксилема), состоящая из трахеид, механических волокон и паренхимы. На поперечном срезе ствола хорошо видны годичные кольца. В коре и древесине многих хвойных имеются смоляные ходы, в которых находятся эфирные масла, смолы и бальзамы, используемые человеком для различных целей.
Игловидные листья, называемые хвоей, у большинства хвойных растений плотные, кожистые, не опадающие ежегодно, однако есть и

Рис. 14. Сосна обыкновенная: 1 – ветка с женскими и мужскими шишками; 2 - семя
Особенности размножения голосеменных рассмотрим на примере сосны обыкновенной (рис.14). Как и все голосеменые - это разноспоровое растение. Взрослое растение сосны является спорофитом. Весной у основания некоторых молодых побегов образуются
собрания зеленовато- желтых мужских шишек - стробилов. На оси мужской шишки расположены микроспорофиллы, на нижней поверхности которых находятся по два микроспорангия (пыльце- вых мешка).
листопадные формы, например, лиственница. Хвоинки покрыты толстым слоем кутикулы, устьица погружены в углубления, заполненные воском. Все это способствует уменьшению испарения воды.
Внутри микроспорангия после редукционного деления образуются гаплоидные микроспоры (пыльцевые зерна). Микроспора имеет две оболочки: наружную - экзину и внутреннюю – интину. В результате отслоения экзины и интины образуются два воздушных мешка, способствующих переносу пыльцы ветром. Ядро микроспоры несколько раз делится, в конечном итоге образуя антеридиальную и вегетативную клетки; первая в дальнейшем дает два спермия, вторая участвует в росте пыльцевой трубки. Таким образом, у сосны мужской заросток (гаметофит) сильно редуцирован и развивается внутри микроспоры.
Антеридий отсутствует.
Женские шишки образуются на верхушках молодых побегов. Женскую шишку нельзя назвать мегастробилом, т.к. она имеет более сложное происхождение. На оси шишки располагаются кроющие (наружные) и семенные (внутренние) чешуи. У основания семенной чешуи находятся два семязачатка (семяпочки). Семязачаток состоит из нуцеллуса и интегумента. Нуцеллус является мегаспорангием. Он срастается с интегументом. Лишь около вершины, обращенной к оси шишки, интегумент имеет отверстие, называемое микропиле или пыльцевход. В мегаспорангии в результате мейоза образуется 4 гаплоидных мегаспоры, из которых только одна развивается дальше. Мегаспора начинает прорастать внутри спорангия и образует женский гаметофит, состоящий из двух архегониев и ткани гаметофита.
При опылении пыльцевое зерно попадает на семяпочку и начинает прорастать, образуя пыльцевую трубку. Сформировавшиеся к тому времени спермии проникают по пыльцевой трубке к архегониям. Один из спермиев сливается с яйцеклеткой, второй погибает. Между процессами опыления и оплодотворения у сосны проходит год. Из образовавшейся зиготы развивается зародыш, состоящий из подвеска, первичного корешка, стебелька и семядолей. Интегумент дает начало семенной кожуре, а из ткани гаметофита образуется гаплоидный эндосперм. Т.е. весь семязачаток превращается в семя. После созревания семян чешуйки шишек расходятся, и семена высыпаются. Зрелое семя сосны имеет крыловидный придаток, способствующий распространению семян (рис. 14, 2).
Хвойные породы играют значительную роль в построении биоценозов, образуя леса на огромных площадях. Основными лесообразующими породами являются сосна обыкновенная, сосна сибирская (кедр), лиственница, пихта, ель. Хвойные используют как строительный материал, топливо; они являются сырьем для огромного
числа веществ, применяемых практически во всех областях народного
хозяйства (смолы, эфирные масла, лекарственные средства и т.д.). Некоторые хвойные культивируются как декоративные растения (кипарисы, туи, можжевельники, тиссы, ели).
Дата добавления: 2018-04-04; просмотров: 3562; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!