Транспорт газів кров’ю. Регуляція дихання.
Дихання – сукупність процесів, які забезпечують надходження в організм кисню, використання його в процесах біологічного окислення та виділення із організму вуглекислого газу.
Існує декілька етапів дихання:
- легенева вентиляція (обмін газів між атмосферним повітрям і альвеолярним),
- газообмін між альвеолярним повітрям і кров’ю (здійснюється завдяки дифузії газів через аерогематичний бар’єр),
- транспорт газів кров’ю,
- газообмін між кров’ю і тканинами,
- клітинне (тканинне) дихання.
Фізіологія вивчає перших чотири етапи (їх об’єднують назвою ”зовнішнє дихання”), біохімія вивчає клітинне дихання (яке називають ”внутрішнім диханням”).
Питання, які ми розглянемо сьогодні, - транспорт газів кров’ю та регуляція дихання.
Кров забезпечує транспорт кисню від легень (де відбувається газообмін між альвеолярним повітрям і кров’ю) до тканин та транспорт вуглекислого газу від тканин (де він утворюється внаслідок процесів біологічного окислення) до легень. Дані гази в крові знаходяться у двох станах: фізично розчиненому та хімічно зв’язаному. Розчинення газів у крові залежить від парціального тиску даного газу (у відповідності до закону Генрі). Нагадую, що парціальний тиск – це тиск газу в суміші газів. У випадку фізичного розчинення газу в рідині (зокрема плазмі крові) ми говоримо про напругу даного газу (зокрема напругу газу в крові).
У 100 мл крові при температурі тіла та нормативних значеннях парціального тиску кисню в альвеолярному повітрі (близько 100 мм рт. ст.) розчиняється лише 0,3 мл О2. Кисень, який розчинився у плазмі крові капілярів малого кола кровообігу, далі надходить в еритроцит (за градієнтом концентрації) і вступає в хімічний зв’язок з гемоглобіном (утворюється оксигемоглобін). У вигляді оксигемоглобіну транспортується близько 20 мл кисню на 100 мл артеріальної крові (тобто 20%)). Слід підкреслити, що одна молекула гемоглобіну може приєднати 4 молекули кисню, що в перерахунку на 1 г гемоглобіну становить 1,34 мл кисню. Таким чином, знаючи кількість гемоглобіну в крові, можна розрахувати, скільки кисню транспортується певним об’ємом крові (100 мл крові).
|
|
|
Киснева ємність крові – кількість кисню, що міститься у 100 мл крові (при кількості гемоглобіну 150 г/л (тобто 15 г на 100 мл крові) - близько 20 мл).
Утворення оксигемоглобіну в капілярах легень залежить від дії декількох факторів:
- напруги кисню (чим більший парціальний тиск кисню в альвеолярному повітрі та напруга кисню в крові, тим більша кількість оксигенованого гемоглобіну), тобто залежність між напругою кисню та кількістю оксигемоглобіну в крові пряма.
|
|
|
- напруги вуглекислого газу (чим більша напруга вуглекислого газу, тим меншим є відсоток оксигенованого гемоглобіну – залежність є оберненою). Слід зауважити, що при високій напрузі кисню у крові (більше 60 мм рт. ст., вуглекислий газ фактично не впливає на оксигенацію гемоглобіну),
- кислотності (залежність обернена). Натомість, залежність між відсотком оксигенованого гемоглобіну та рН – пряма,
- температури (залежність обернена),
- кількості 2,3-дифосфогліцерату (проміжний продукт метаболізму глюкози, який утворюється в еритроцитах в процесі гліколіза, залежність обернена).
При високих напругах кисню всі перераховані фактори майже не впливають на процес зв’язування кисню з гемоглобіном.
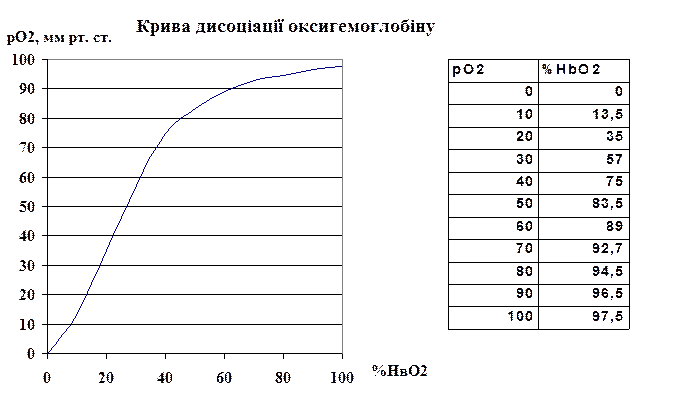
Залежність між напругою кисню і кількістю оксигенованого гемоглобіну, як ми вже згадували, пряма, проте не лінійна, вона виражається s-подібною кривою (s-подібна крива дисоціації оксигемоглобіну).
Ліва частина кривої відтворює процеси в тканинах, коли при низькій напрузі кисню (<60 мм рт. ст.) усі фактори призводять до зниження відсотка оксигемоглобіну (кисень від’єднується від оксигемоглобіну і дифундує у тканини). Права частина кривої відображає процеси в легенях, коли при високих напругах кисню (60 – 100 мм рт. ст.) на утворення оксигемоглобіну усі фактори (СО2, температура, рН) практично не впливають, крива стає практично горизонтальною, майже весь гемоглобін зв’язаний з киснем (відсоток оксигемоглобіну – 96-98%).
|
|
|
У капілярах великого кола кровообігу кисень (внаслідок різниці напруг газу в крові і клітинах) дифундує до клітин, де використовується в окисних процесах. Клітини у стані спокою споживають близько 30-40% кисню артеріальної крові, під час фізичного навантаження – до 60 - 70%. Частка кисню, що споживається тканинами, називається коефіцієнтом утилізації кисню (КУК):
КУК= [O2] в артеріальній крові – [O2] у венозній крові · 100% = 20% - 14% · 100% = 30%
[О2] в артеріальній крові 20%
У венозній крові частина гемоглобіну залишається в оксигенованій формі (близько 15 мл на 100 мл крові), фізично розчиненого кисню – близько 0,12 мл (також на 100 мл крові).
Вуглекислий газ, який утворився в клітинах у процесі біологічного окислення, дифундує із клітин в плазму крові капілярів великого кола кровообігу (за градієнтом концентрації). Фізично розчиненого СО2 – 2,5 – 2,6 мл на 100 мл крові. У той же час із 100 мл венозної крові можна виділити 58 мл вуглекислого газу. Отже, більша частина СО2 знаходиться в хімічно зв’язаному стані – у вигляді карбгемоглобіну (близько 4,5 мл на 100 крові) та солей вугільної кислоти (бі- або гідрокарбонатів) – до 51 мл на 100 крові. Відбувається це внаслідок того, що вуглекислий газ постійно шляхом дифузії (за градієнтом концентрації) переходить із плазми крові капілярів великого кола кровообігу в еритроцити, де сполучається з гемоглобіном (утворюється карбгемоглобін), а також (під дією ферменту карбоангідрази) сполучається з водою (з утворенням вугільної кислоти). Остання швидко дисоціює на протон і гідрокарбонат-аніон. Протон приєднується до гемоглобіну (працює гемоглобіновий буфер), а гідрокарбонат виходить у плазму крові за градієнтом концентрації, а на його місце (для стабілізації осмотичного тиску в еритроциті та заряду мембрани) із плазми крові заходить іон хлору. Останній процес має назву хлоридне зрушення. Коли кров надходить до капілярів малого кола кровообігу, відбуваються зворотні процеси.
|
|
|
Під регуляцією дихання розуміють пристосування зовнішнього дихання до потреб організму зі стабілізацією напруг кисню та вуглекислого газу в крові та рН крові. Ритм та глибина дихання задаються та регулюються центральними нервовими структурами, розташованими у людини в декількох відділах мозку і умовно об’єднаних в поняття “дихальні центри”. Під впливом на ці центри рефлекторних та гуморальних стимулів відбувається пристосування дихання до змін умов зовнішнього середовища та стану самого організму.
Виділяють спинальний дихальний центр, представлений мотонейронами передніх рогів спинного мозку, які забезпечують іннервацію дихальних м’язів (зокрема, це мотонейрони III та IV шийних сегментів, що забезпечують іннервацію діафрагми; мотонейрони грудних сегментів, які забезпечують іннервацію міжреберних м’язів та м’язів живота ). При перерізці спинного мозку на рівні нижніх шийних сегментів дихальна активністьдіафрагми зберігається, а міжреберних м’язів – припиняється. При перерізці спинного мозку на рівні верхніх шийних сегментів дихання повністю зникає. Таким чином, за відсутності імпульсів від вищерозташованих дихальних центрів активність спинальних мотонейронів не забезпечує адекватного дихання.
Бульбопонтинний дихальний центр (згідно своєї назви) знаходиться у довгастому мозку та варолієвому мосту, представлений дихальними нейронами, імпульсна активність яких змінюється у відповідності до фаз дихального циклу (тобто фази вдиху і видиху). У довгастому мозку вказані нейрони сконцентровані у вигляді дорзального та вентрального ядер. Нейрони дорзального ядра інспіраторні (inspiratio – вдих), бульбоспинальні, їх аксони утворюють синапси з мотонейронами шийних сегментів спинного мозку (ядрами діафрагмального нерва ). Вентральне ядро довгастого мозку містить як інспіраторні, так і експіраторні (expiratio – видих) нейрони, які пов’язані з мотонейронами міжреберних м’язів та м’язів черевного преса. У мосту знаходиться пневмотаксичний та апнейстичний дихальні центри, які модулюють активність нейронів вентрального та дорзального ядер довгастого мозку, регулюючи, відповідно, частоту і глибину дихання. Пневмотаксичний центр розташований у верхній третині моста і представлений нейронами nucleus parabrachialis, апнейстичний центр локалізований у середній та нижній частині моста, представлений ретикулярними нейронами. При перерізці моста нижче чотиригорбикового тіла (та одночасній перерізці блукаючих нервів) дихання зберігається, проте частота дихання різко зменшується, вдих подовжується (виникає так зване ”апнейстичне дихання”). При перерізці між мостом та довгастим мозком виникає гаспінг-дихання, для якого характерні швидкі короткі вдихи та тривалі експіраторні паузи.
Гіпоталамус причетний до регуляції дихання під час поведінкових актів, він узгоджує діяльність вегетативних центрів з дихальними.
Поведінкова регуляція дихання здійснюється корою головного мозку. Складні зміни дихання відбуваються під час співів, мови, супроводжують психічні процеси (мислення, увага, емоції).
Інформація про параметри зовнішнього дихання надходить до бульбопонтинного дихального центру та вищерозташованих центрів ЦНС від механорецепторів легень та хеморецептивних зон, а також від деяких інших полімодальних рецепторів.
Хеморецептори забезпечують аферентну сигналізацію про газовий склад внутрішнього середовища організму. Виділяють центральні та периферичні хеморецептивні зони. Центральні хеморецептори розташовані на вентролатеральній поверхні довгастого мозку біля виходу IX та X пар черепномозкових нервів на глибині 0,2-0,4 мм (рецептивні поля М і L). Адекватним подразником для них є концентрація іонів водню в спинномозковій рідині та міжклітинній рідині мозку (так, зниження рН спинномозкової рідини на 0,01 супроводжується збільшенням вентиляції легень на 4 л/хв.). У свою чергу концентрація іонів водню залежить від напруги СО2 в артеріальній крові (СО2 проходить через гематоенцефалічний бар’єр, СО2+Н2О→Н2СО3→ Н+ +НСО3-). Роль напруги СО2 в регуляції дихання доведена Л.Фредеріком у досліді з перехресним кровообігом у собак (1890, 1901). У двох собак, які знаходились під наркозом, перерізали і перехресно з’єднали сонні артерії і окремо яремні вени. Після перев’язки хребцевих артерій голова кожної собаки отримувала кров від тулуба іншої собаки. Якщо у першої собаки перетиснути трахею (штучна асфіксія, припинення вентиляції легень), то у другої собаки розвивається гіпервентиляція (що пов’язано із накопиченням СО2 в крові першої собаки та надходженням крові, збагаченої вуглекислим газом, до голови другої собаки, дихальний центр якої збуджується). Внаслідок гіпервентиляції напруга СО2 в крові другої собаки знижувалась; кров, збіднена вуглекислим газом, надходила до голови першої собаки, дихальний центр першої собаки пригнічувався, виникало апное, дихальні рухи припинялися.
Периферичні хеморецептори розташовані в каротидних тільцях, які знаходяться в ділянці біфуркації загальних сонних артерій, та в аортальних тільцях, які є на верхній та нижній поверхнях дуги аорти та в ділянці правого плечеголовного стовбура. Адекватним подразником для них є величина напруги кисню в артеріальній крові, а також підвищення напруги вуглекислого газу та зниження рН крові. Імпульсація від хеморецепторів каротидних тілець надходить до дихального центру (зокрема, до дорсальної дихальної групи нейронів довгастого мозку) по нерву Герінга (гілка IX пари черепномозкових нервів), а від хеморецепторів аортальних тілець – по чутливим волокнам n.vagus. Сигнали, що надходять до довгастого мозку від центральних та периферичних хеморецепторів, є необхідною умовою періодичної активності дихального центру.
Механорецептори легень (їх ще називають рецепторами розтягування легень) локалізовані в гладком’язовому шарі стінок трахеобронхіального дерева, вони оцінюють ступінь розтягування альвеол та дихальних шляхів. Імпульси від цих рецепторів надходять до дихального центру по чутливим волокнам блукаючого нерва, сприяючи зміні дихальних фаз (зміні вдиху на видих). Вплив механорецепторів на регуляцію дихання доведено в дослідах Е. Герінга та І. Брейєра (1868). Дані дослідники встановили, що збільшення об’єму легень викликає наступні рефлекторні ефекти:
- дослід 1. Якщо під час вдиху собаці додатково в легені вдувати повітря, вдих достроково припиняється (виникає інспіраторно-гальмівний ефект),
- дослід 2. Вдування повітря в легені собаки під час видиху викликає подовження експірації (експіраторно-полегшуючий ефект),
- дослід 3. Досить сильне роздування легень викликає короткочасне, однак сильне збудження інспіраторних м’язів, виникає судомний вдих (це парадоксальний ефект Хеда).
Зменшення об’єму легень обумовлює посилення інспіраторної активності і прикорочення видиху, тобто сприяє виникненню наступного вдиху (рефлекс на спадіння легень).
До полімодальних рецепторів, причетних до регуляції дихання, відносять:
- іритантні рецептори (розташовані в епітеліальному та субепітеліальному шарах всіх повітроносних шляхів, реагують на різкі зміни об’єму легень, а також на пил, холодне повітря, їдкі гази, тютюновий дим, гістамін. Їх аферентні волокна ідуть у складі блукаючого нерва. Подразнення іритантних рецепторів через блукаючий нерв може призвести до бронхоспазму (що часто виникає при бронхіальній астмі);
- J-рецептори (юкстакапілярні) (локалізовані в інтерстиції легень поблизу капілярів альвеол, дають початок немієлінізованим С-волокнам, які проходять у складі блукаючого нерва, чутливі до збільшення об’єму інтерстиціальної рідини в легеневій тканині, ряду хімічних речовин (нікотин, гістамін, простагландини);
- рецептори верхніх дихальних шляхів (чутливі до слизу, чужорідних тіл, хімічних подразників, є джерелом рефлексів захисного характеру – кашлю, чхання, бронхоспазму).
Окремою групою рецепторів є пропріорецептори дихальних м’язів. Вони контролюють відповідність скорочень дихальних м’язів тому завданню, яке м’язи отримали від дихального центру.
Дата добавления: 2015-12-21; просмотров: 28; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!