Синтез жирных кислот.
Обмен липидов.
В среднем организм человека получает около 80 г. жиров растительного и животного происхождения в сутки. Жиры имеют важное энергетическое значение для организма, при расщеплении 1 г жира выделяется около 38,9 кДж (1 г белков или углеводов - 17,2 кДж). Жиры являются растворителями витаминов, источником незаменимых жирных кислот, фосфолипидов, стеринов.
Жиры пищи не перевариваются слюной, в желудке отмечается частичное разрушение липопротеидных комплексов мембран. У человека и млекопитающих расщепление жиров происходит преимущественно в верхних отделах тонкого кишечника.
В двенадцатиперстную кишку поступает желчь и панкреатический сок, необходимые для эмульгирования и переваривания жиров. Происходит нейтрализация желудочного сока бикарбонатами панкреатического скока. Пузырьки углекислого газа способствуют перемешиванию кашицы с соками. В соке поджелудочной железы содержатся липазы, имеющие оптимум рН - 6 (в присутствии желчи он смещается с 8 до 6).
Желчь содержит желчные кислоты, у человека основная кислота - холевая (дезоксихолевая, литохолевая и хенодезоксихолевая) и коньюгированные кислоты - гликохолевую и таурохолевую.
NН2-СН2-СООН - глицин
NН2-СН2-SО3Н - таурин
Они способствуют эмульгированию так же как и мыла.
Под действием липаз от триацилглицерина отщепляются сначала быстро жирные кислоты в положении 1 и 3, в в положении 2 - медленно.
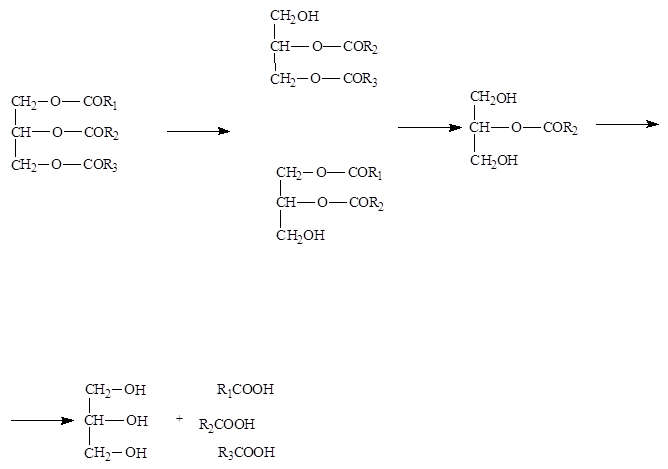
Расщепление фосфолипидов идет под действием фосфолипаз А1, А2, С и D и лизофосфолипаз. Стериды расщепляются под действием холестераз поджелудочной железы с образованием холестерола и жирных кислот. В результате гидролиза липидов образуются анионы жирных кислот, моно, ди и триацилглицерины, эмульгированные мылами, глицерин, холин, этаноламин, холестерол. Всасываются все продукты расщепления и в небольшой степени (3-10%) нерасщепленные жиры.
Глицерин и жирные кислоты с количеством углеродных атомов до 10 всасываются и поступают в печень через портальную систему кровообращения. Жирные кислоты, имеющие более 10 атомов углерода в цепи и моноацилглицерины образуют мицеллы с желчными кислотами и мылами и могут проходить через водный слой слизистой кишечника. Жирные кислоты образуют растворимые комплексы с гликохолевой и таурохолевой кислотой - холеиновые кислоты, которые легко всасываются в эпителии кишечника.
Из стенки кишечника желчные кислоты всасываются в кровь, поступают в печень и опять возвращаются в желчь. Часть желчных кислот (0,2-0,5 г) в сутки выводится с калом.
Лучше перевариваются и всасываются липиды, находящиеся в жидком состоянии при температуре тела. При более высокой температуре плавления жиры плохо перевариваются и всасываются.
Фосфорная кислота всасывается в виде солей, а азотистые основания всасываются при участии нуклеотидов.
Среди стероидов только холестерин легко проходит через стенку кишечника. Также проникает витамин D и некоторые стероидные гормоны.
Большая часть продуктов переваривания в клетках кишечника вновь превращается в триацилглицерины. Ресинтезированные триа-
цилглицерины, фосфолипиды, холестерин и его эфиры в стенках эпителия кишечника соединяются с небольшим количеством белка и образуют хиломикроны (100-500 нм). Они проникают в лимфатическую систему кишечника, из нее через грудной лимфатический проток в кровяное русло.
Хиломикроны транспортируют экзогенные липиды в печень и жировую ткань. Они легко диффундируют из крови в межклеточные пространства печени. Гидролиз триацилглицеринов может происходить внутри клеток печени и на их поверхности. В клетки жировой ткани хиломикроны проникать не могут и гидролиз идет на поверхности эндотелия капилляров жировой ткани под действием липопротеинлипазы.
Часто образовавшихся жирных кислот проникает в клетки и запасается в виде липидов, другая часть связывается с альбуминами сыворотки крови и разносится в органы и ткани. Резервные липиды на 99% являются триацилглицеринами. Они непрерывно мобилизуются и одновременно такое же количество липидов запасается.
Метаболизм резервных липидов начинается с гидролиза с участием липопротеинлипазы крови или жировой ткани. Жирные кислоты связываются с альбумином крови и переносятся в печень, где происходят их главные превращения.
Глицерин может включаться в гликолиз с предварительным образованием глицерин-3-фосфата или включаться в глюконеогенез.
глицеролкиназа дегидрогеназа
Глицерин + АТФ --------------® Глицерол -3- Ф --------------®
НАД+ НАДН
--------® Глюконеогенез
---® Триозофосфаты |
----------® Гликолиз
Окисление жирных кислот в клетках происходит в митохондриях при участии мультиферментного комплекса путем и-окисления. АТФ стимулирует образование тиоэфирной связи между карбоксилом жирной кислоты и сульфгидрильной группой кофермента А под дейс-
твием ацил-КоА-синтетазы (тиокиназа жирных кислот).
R-СО-О - + АТФ + Н-S-КоА --------® R-СО-S-КоА + АМФ + Н4Р2О7
Жирные кислоты активируются на наружной митохондриальной мембране, а окисляются в митохондриальном матриксе. Ацил-КоА переносится карнитином. 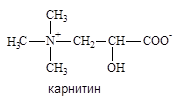
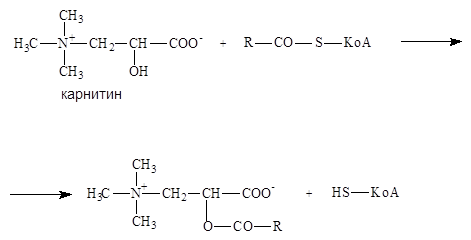
После прохождения через мембрану идет обратная реакция (карнитин-ацил-КоА-трансфераза).
Насыщенный Ацил-КоА распадается в результате повторяющейся последовательности из четырех реакций:
1. Окисление с участием флавинадениндинуклеотида.
2. Гидратация.
3. Окисление с участием НАД.
4. Тиолиз с участием кофермента А.

3. Н О Н
| | 1-3-гидроксиацил-КоА-
R-СН2-С-С-СО-S-КоА + НАД+ --------------------®
| | дегидрогеназа
Н Н
-----® R-CН2-СО-СН2-СО-S-КоА + НАДН + Н+
3-кетотиолаза
4. R-CН2-СО-СН2-СО-S-КоА + НS-КоА ---------------®
---® R-СН2-СО-S-КоА + Н3С-СО-S-КоА
При этом цепь укорачивается на два углеродных атома и происходит генерирование ФАДН2 и НАДН и ацетил-КоА. Первые три реакции расщепления жирных кислот сходны с последними реакциями цикла ТКК. Укороченный ацил-КоА подвергается далее следующему циклу окисления.
В каждом цикле реакций образуется по одной молекуле ФАДН2, НАДН и ацетил-КоА.
Сn-ацил-КоА + ФАД + НАД+ + Н2О + КоА-S-Н -----------®
---® Сn-2-ацил-КоА + ФАДН2 + НАДН + Ац-КоА + Н+
При окислении жирных кислот выделяется большое количество энергии.
Пальмитоил-КоА + 7 ФАД + 7 НАД+ + 7 НS-КоА + 7Н2О ---------®
---® 8 Ац-КоА + 7 ФАДН2 + 7 НАДН + 7 Н+
Пальмитиновая кислота окисляется за 7 циклов. В дыхательной цепи НАДН образует 3АТФ, ФАДН2 - 2АТФ, Ац-КоА - 12 АТФ. Итого образуется: 7х3 + 7х2 + 8х12 = 131 АТФ. На активацию молекулы пальмитиновой кислоты была затрачена 1АТФ. В результате окисления пальмитиновой кислоты образуется 130 молекул АТФ. К.п.д. процесса около 45%, что близко к гликолизу, ЦТК, окислительному фосфорилированию.
Жирные кислоты, имеющие нечетное число атомов углерода на последнем этапе дают пропионил-КоА. Активированный трехуглеродный фрагмент пропионил-КоА включается в ЦТК после его превращения в сукцинил-КоА.
Окисление ненасыщенных кислот происходит также, как и насыщенных, но после удаления нескольких Ац-КоА требуется еще фермент - ^-3,4-цис-^2,3-трансеноил-КоА-изомераза, который осуществляет переход двойной связи в положение 2,3 и изменение конфигурации.
Для окисления полиненасыщенных кислот требуется еще один дополнительный фермент - 3-гидроксибутирил-КоА-эпимераза, кото-
рый переводит D-изомер в L-изомер, легко окисляемый специальной
дегидрогеназой. Наличие изомеразы и эпимеразы обеспечивает возможность полного окисления всех ненасыщенных жирных кислот, содержащихся в липидах.
Ацетил-КоА, образовавшийся при окислении жирных кислот включается в цикл ТКК в условиях, когда расщепление жиров и углеводов сбалансировано. Включение Ац-КоА в цикл зависит от доступности оксалоацетата для образования цитрата. Если расщепление жиров преобладает, то Ац-КоА распадается по другому пути.
В случае голодания или сахарного диабета оксалоацетат расходуется на образование глюкозы и не может конденсироваться с Ац-КоА. В этих условиях образуются кетоновые тела: ацетоацетат, 3-гидроксибутират и ацетон.
2Ац-КоА + Н2О ----® СН3-СО-СН2-СОО- + 2 НS-КоА + Н+
При восстановлении ацетоацетата образуется 3-гидроксибутират. Ацетоацетат медленно спонтанно декарбоксилируется в ацетон.
ацетоацетил-КоА-тиолаза
2 СН3-СО-S-КоА -----------------------® СН3-СО-СН2-СО-S-КоА + НS-КоА
b-окси-b -метил-глутарил-Ко-А-лиаза
СН3-СО-СН2-СО-S-КоА + СН3-СО-S-КоА --------------------------®
ОН
|
-® НООС-СН2-С-СН2-СО-S-КоА
|
СН3
ОН
|
НООС-СН2-С-СН2-СО-S-КоА ---------------® СН3-СО-СН2-СООН + Ац-КоА
|
СН3
гидроксибутиратдегидрогеназа ОН
СН3-СО-СН2-СООН + НАДН2 ----------------------------® |
СН3-СН-СН2-СООН
СН3-СО-СН2-СООН -----------® СН3-СО-СН3 + СО2
Ацетоацетат и гидроксибутират имеют физиологическое значение. В норме они являются дыхательным топливом и количественно важными источниками энергии. их потребляют сердечная мышца и корковое вещество надпочечников. Глюкозу употребляет мозг. Он при голодании и сахарном диабете адаптируется к использованию ацетоацетата.
Соотношение кетоновых тел в кровотоке колеблется: если в печени много гликогена, то образуется больше 3_гидроксибутирата, если гликогена мало, преобладает ацетоацетат.
Животные неспособны превращать жирные кислоты в глюкозу. Ац-КоА на может превращаться в пируват или оксалоацетат. Оксалоацетат регенерируется, а не образуется заново в цикле ТКК. Растения имеют два дополнительных фермента и способны превращать жиры в глюкозу.
Синтез жирных кислот.
Синтез жирных кислот разобщен с их распадом.
1. Синтез идет в цитозоле.
2. Промежуточные продукты синтеза жирных кислот связаны с ацилпереносящим белком.
3. Ферменты синтеза организованы в мультиферментный комплекс - синтетаза жирных кислот.
4. Растущая цепь удлиняется путем присоединения двууглеродных остатков.
5. Роль восстановителя играет НАДФН.
6. Синтез заканчивается на пальмиате.
Синтез жирных кислот начинается с карбоксилирования ацетил-КоА в малонил-КоА под действием ацетил-КоА-карбоксилазы. В качестве простетической группы она содержит витамин Н -биотин.
Образующийся в митохондриях ацетил-КоА взаимодействует с оксалоацетатом с образованием цитрата, который свободно проникает в цитоплазму клетки, где он расщепляется до Ац-КоА и оксалоацетата. Цитрат выступает в роли переносчика ацетильного радикала.
Н3С-СО-S-КоА + НСО3- + АТФ -----® -ООС-СН2-СО-S-КоА + АДФ +Н3РО4
Эта реакция является лимитирующей для всего процесса синтеза жирных кислот и может ускоряться цитратом, изоцитратом и кетоглутаратом.
Ацетил-КоА и малонил-КоА вступают в реакцию с ацилпереносящим белком. Он состоит из одной полипептидной цепи, содержащей 77 аминокислотных остатков и является гигантской простетической группой - макро-КоА. Связь осуществляется с сульфгидрильным концом фосфопантотеновой кислоты. Этот компонент является частью Ко-А и связан с остатком серина в АПБ.
ОН СН3 О-
| | |
Н-S-(СН2)-NН-СО-(СН2)2-NН-СО-СН-С-СН2-О-РО-О-СН2-Ser-АПБ
|
СН3
Синтез начинается с образования ацетил-АПБ и малонил-АПБ. Жирные кислоты с нечетным числом атомов углерода начинаются с пропионил-АПБ.
СН3-СО-S-КоА + НS-АПБ ----® СН3-СО-S-АПБ + НS-КоА
НООС-СН2-СО-S-КоА + НS-АПБ ----® НООС-СН2-СО-S-АПБ + НS-КоА
Ацетил-АПБ и малонил-АПБ взаимодействуют с образованием ацетоацетил-АПБ под действием 3-кетоацил-АПБ-синтетазы.
СН3-СО-S-АПБ + НООС-СН2-СО-S-АПБ ----® СН3-СО-СН2-СО-S-АПБ +НS-АПБ + СО2
Синтез из двух ацетильных остатков невыгоден энергетически. Присоединение малонила с одновременным декарбоксилированием идет с выделением энергии и поэтому протекает активно не требуя дополнительных условий.
Все жирные кислоты с четным числом атомов углерода происходят из Ацетил-КоА.
3-кетоацил-АПБ-редуктаза
1. СН3-СО-СН2-СО-S-АПБ + НАДФН + Н+-------------------------®
----® СН3-СН-СН2-СО-S-АПБ + НАДФ+
|
ОН
3-гидроксиацил-АПБ-дегидратаза
2. СН3-СН-СН2-СО-S-АПБ - Н2О ----------------® СН3-СН=СН-СО-S-АПБ (кротонил-АПБ)
|
ОН
еноил-АПБ-редуктаза
3. СН3-СН=СН-СО-S-АПБ + НАДФН + Н+ -------------®СН3-СН2-СН2-СО-S-АПБ + НАДФ+
(бутирил-АПБ)
Следующие за конденсацией восстановление, дегидратация и второе восстановление приводят к образованию бутирил-АПБ, содержащего четыре углеродных атома.
Во втором цикле бутирил-АПБ конденсируется с малонил-АПБ с образованием шестиуглеродного фрагмента, затем идет образование восьмиуглеродного фрагмента и т.д. до шестнадцатиуглеродного соединения (пальмитоил-АПБ).
Пальмитоил-АПБ гидролизуется с образованием пальмитиновой кислоты. Суммарное уравнение реакции:
Ац-КоА + 7 Мал-КоА + 14 НАДФН + 7 Н+® пальмиат + 7СО2 +14 НАДФ+ + 8 НS-КоА+ +6Н2О
7 Ац-КоА + 7СО2 + 7АТФ ------® 7 Мал-КоА + 7АДФ + 7Н3РО4 + 7Н+
8 Ац-КоА + 7АТФ + 14НАДФН -----® пальмиат + 14НАДФ+ + + 8НS-КоА + 6Н2О + 7АДФ + Н3РО4
Человек и высшие животные пополняют запас жирных кислот синтезом пальмитиновой кислоты и поступлением их с пищей. Пальмитиновая и стеариновая кислоты являются источником синтеза мононенасыщенных кислот. Полиненасыщенные кислоты в организме не синтезируются, являются незаменимыми и должны поступать с пищей.
Синтез триацилглицеринов идет из производных жирных кислот через фосфатидную кислоту в печени и жировой ткани. Глицерин фосфорилируется глицеролкиназой и АТФ.
СН2ОН СН2-О-СО-R
| |
СН-ОН + 2 R-СО-S-КоА -------® СН-О-СО-R + 2 НS-КоА
| |
СН2-О-Р СН2-О-Р
При гидролизе фосфатидной кислоты образуется 1,2-диацилглицерин, который реагирует с еще одним Ацил-КоА и образует глицерид.
В слизистой кишечника триацилглицерины синтезируются из свободных жирных кислот, моно- и диацилглицеринов. Остаток жирной кислоты переносится в виде Ацил-КоА. Глицерофосфолипиды синтезируются из фосфатидной кислоты.
Фосфатидная кислота
|
| + УТФ
УДФ-диацилглицерин
--------------------------------------------------------------------------
| | |
+ серин | + инозит | + глицерофосфат |
| | |
Фосфадитилсерин Фосфадитилинозит Фосфадитил-
| глицерол-Ф
|-СО2 |
Фосфадитилэтаноламин | -Н3РО4
| Фосфадитилглицерин
+ 2СН3 | |
| + фосфадитил- | -глице-
Фосфадитилхолин глицерин | рин
|
Кардиолипин
Терпены и стерины, имеющие в основе строения изопрен, синтезируются из мевалоновой кислоты. Молекула холестерина также образуется через мевалоновую кислоту целиком из ацетильных остатков ацетил-КоА. Один из промежуточных продуктов - и-гидрокси-и-метилглутарил-КоА образуется и при синтезе кетоновых тел.
СН3
|
3 Ац-КоА ------® НООС-СН2-С-СН2-СО-S-КоА + 2 НАДФН -------®
|
ОН
СН3
|
---® НООС-СН2-С-СН2-СН2ОН + 2 НАДФ+ + 3 НS-КоА
|
ОН мевалоновая кислота
Мевалоновая кислота далеее подвергается ряду превращений,в ходе которых отщепляется углекислый газ, а пятиуглеродные части шести молекул мевалоновой кислоты конденсируются, образуя сквален:
СН3 СН3 СН3 СН3 Н3С
| | | | |
Н3С-С=СН-СН2-СН2-С=СН-СН2-СН2-С=СН-СН2-СН2-СН=С-СН2-СН2-СН-С
|
Н2С
|
Н3С-С=СН-СН2
|
СН3
Сквален затем превращается в ланостерин, из которого в несколько стадий образуется холестерин. Около 80% холестерина образуется в печени, 10 - в тонком кишечнике, 5 - в клетках кожи. За сутки в организме человека синтезируется до 1г холестерина. Скорость синтеза регулируется по механизму обратной отрицательной связи. Основным местом регуляции является образование мевалоновой кислоты. При содержании в пище 2-3 г холестерина в сутки синтез собственного холестерина полностью прекращается. Холестерин является источником синтеза многих важных веществ:
Холестерин
Желчные Стероиды
кислоты растений
Половые Стерины
гормоны кала
Кортикостероиды Витамин D
В организме взрослого человека в сутки обновляется 1,3 г холестерина. С пищей поступает около 0,5 г холестерина в сутки. Всасывается около 30% холестерина. Желчь поставляет в кишечник
1,5-2,0 г холестерина. Выведение холестерина происходит путем окисления его в желчные кислоты, которые удаляются с калом (0,5
г), также сам холестерин удаляется с калом. При нарушении баланса холестерина возникает гиперхолестериномия. Она учеличивает вероятность заболеваний атеросклерозом и желче-каменной болезнью.
Если количество употребляемых углеводов превышает необходимое для возобновления запасов гликогена, то часть углеводов превращается в жиры. Глюкоза служит источником ацетил-КоА, из которого синтезируются жирные кислоты. Глицерофосфат образуется при восстановлении диоксиацетонфосфата.
СН2ОН СН2ОН
| |
С=О + НАДН + Н+ -----------® СН-ОН + НАД+
| |
СН2-О-Р СН2О-Р
Из глюкозы образуется все, что необходимо для синтеза жиров. Синтез идет через фосфатидную кислоту.
Синтез жиров из углеводов наиболее активно происходит в печени, менее активно в жировой ткани.
У нормально упитанного человека жиры составляют 15% массы тела. При полном голодании этот запас расходуется 5-7 недель. При нормальном питании количество жира в организме не изменяется. Жиры обновляются постоянно.
Наиболее частой причиной ожирения является несоответствие между количеством потребляемой пищи и энергетическими затратами организма. Это чрезмерное потребление пищи и гиподинамия, и особенно сочетание этих факторов.
При ежедневном употреблении 3 г лишней пищи за 10 лет образуется избыточная масса тела в 10 кг.
Обмен липидов регулирует эндокринная система. bl-13
Дата добавления: 2015-12-16; просмотров: 54; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!