Майлардың ыдырауы.
Ағзада майлардын биологиялық маңызы алуан түрлі, олардың негізгілері:
1) майлар мен май тәрізді заттар ағза үшін қуат көзі болып табьшады. Тәулігіне жүмсалған куат кезінің 25-30% пайызы липидтер аркьшы өтеледі: 1 г май тотыққанда 38,9 кДж (9,3 ккал) қуат бөлінеді;
2) липидтер жасуша цитоплазмасы мен мембрананыц күрамына кіреді. Олар көмірсулармен қосылып жүйке үлпаларының күрамыи құрайды;
3)ішкі мүшелердің айналасында, шарбыда, шелде кор ретінде жинақталған липидтер сол мүшелерді соғылудан, үйкелуден сақтайды; 4) майлар майда еритін витамиңдерді (АД,Е және К) ерітіп, оларды ағзаға таратады;
5) липидтер ағзадағы жылудын реттелуіне (сакталуына) қатысады, теріні майлап, кұрғап кетуден сақгайды.
Барлық липидтер құрамының ерекшелігіне байланысты қарапайым және күрделі болып екі топқа бөлінеді:

Тамақтың қүрамындағы стеридтер холиэстеразаның әсерінен холестерин және жоғары молекулалы май қышқылдарына ыдырайды.
Қорытылудың әсерінен түзілген липидтердің суда жақсы еритін өнімдері: глицерин, фосфор қышқылы, азотты негіздер, моноглицеридтер ішек аркылы оңай сіңеді. Ал суда ерімейтін өнімдердің (май қышқылы, холестерин т.б.) сіңуі үшін парлы ет қышқылдарының маңызы зор. Сіңгеннен сон глицерин мен май кышкыпдары косылып ағзаға тән үшглицеридтер мен фосфолипидтер пайда болады.
|
|
|
Пайда болған май мен май тәрізді заттар белокпен косылып хиломикрондар (мөлшері 100-ден 5000 нм дейін) түзеді. Олар лимфаға сіңіп, одан соң қанға өтеді. Қан аркылы ағзаның барлык мүшелеріне тарап оның бірталай мөлшері тері астындағы шелде, шарбыда т.б. кор ретінде жинақталады.
Глицерин тотығуы оның АҮФ-ның куат әсерінен фосфорлануынан
басталып, ол фосфоглицератқа айналады. Соңғысынан 3-фосфоглицератты альдегид түзіліп ары қарайғы жолы көмірсулардың тотығуымен бірдей жүреді (26-кесте). Сонымен қатар бір молекула глицерин тотыққанда жаңадан 22 молекула АҮФ пайда болады.

3. Май қышқылының тотығуы негізінен бауыр жасуша митохондрийінде өтеді. Негізінен құрамында жұп көміртегі атомы бар жоғары молекулалы майлы кышқылдар тотығады. Майлы қышкылдар АҮФ-ның қуатының әсерінен белсенді майлы қышқылға айналады. Соңғысы цитоплазмадағы карнитиннің қатысуымен митохондрийге етіп, онда β - тотығуға ұшырайды. Майлы қышқылдардың β -тотығуын 1904 жылы бірінші рет Ф.Кнооп дәлелдеген. β -тотығуы деп аталатын себебі, майлы қышқылдар тотыға келіп, әрқашан 2 көміртегі атомына қысқарады (карбоксил тобынан кейінгі β- жағдайында тұрған екінші көміртегі атомы) (27-кесте).
|
|
|
Майлы қышкылдардың β -тотығуының бір циклында 5 молекула АҮФ түзіліп, пайда болған белсенді сірке қышкылы Кребс цикліңде тотығып 12 молекула АҮФ береді. Сондықтан бір молекула үш стеарин толық тотыққ анда 463 молекула тузіліп, қуатының тиімділігі 63% пайыздай болады.
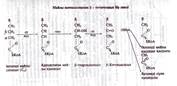
Қалыпты жағдайда май қышқылынын β -тотығуы арқылы бауырда кетоденелер: ацетон, ацетонсірке қышкылы, β -гидромайқышқылы түзіледі. Қандағы кетоденелердің мөлшері ммоль.л-1 10-50 мг/л, несеппен тәулігіне 40 мг шамасындай кетоденелер сыртқа бөлінеді. Бүл мөлшерден артык кетоденелердің пайда болуы:
1) ағзаға азык-түлікпен енгізілген көмірсулардың мөлшері азайғанда;
2) ашыққанда;
3) ағзаға кейбір аминқышқылдар (лейцин, фенилаланин) көп түссе;
|
|
|
4) ұзақка созылған дене еңбегінде;
5) көмірсулардың жэне майлардың алмасуы бүзылса (диабет).
Бұл кезде несеппен бөлінетін кетоденелердің мөлшері 10-нан 50г
дейін өсуі мүмкін.
4. Жогары май қышқылдэрының биосинтезі
Қаныққан жогары май қышқылдарының синтеэі барлық тірі ор-ганиэмде, көбінесе май тканьдерінде, сүт бездеріңдө және бауырда журеді. Май қыщқылдарының тотыгу процесеі клетканың митохондрисында журсе, синтезі цитоплазмада журеді. Сннтез журетін рөакция-лардың жалпы теңдеуі тамендегідей:
Ацетид-КоА + 7 Малонил -S- СоА + 14 НАДФН+→Пальмитин қышқылы + 7СО2 + 14 НАДФ+ + 6 Н2О;
Май қышқылдары синтезінің кейбір ерекшеліқтері:
1. Синтеэ цитоплазмадагы эндоплазмалық торда журеді.
2. Синтеэ ушін ацетил-КоА-дан туэілетін малонил-КоА жумсалады.
3. Ацетил-КоА - синтез үшін тек синтездік реакциялардың басталуы-на гана қажет, ягни "үйытқысы" болыл табылады /эатравка/.
4. Май қышқылдарының синтезіндері аралық заттарды тотықсызданды-ру ушін, глюкоэаның пентозофосфаттық жолмен ыдырауы кезіндө тузілетін НАДФН+ жүмсалады.
|
|
|
5. Малонил-КоА-ның қатысуымен журөтін май қышкылдарының синтеэі пальмитатсинтетаза ферменттік қомплексінің қатысумен журетін циклді лроцеес. Малонил-КоА-ның синтеэі үшін ацетил-КоА қажет, Ацетил-КоА клетканың мнтохондриіңде жинақтаяган. Осы. Аце-
тил-КоА-ның митохондрийден цитоплазмаға өткізудің екі меха-ниэмі бар: I/ Карңитиннің және ацетил~КоА-карнитин~трансфе раэа ферментінің көмегімен митохондрийден цитоплазмаға өтуі;
Ацетил-КоА + карнитин митох.
'цитоплазма. Ацетил - карнитин
.
2/ Цитраттың митохондрийден өтіп, АТФ - цитратлиаза ферментінің қатысуымен ыдырау механизмі.
Цитрат +АТФ+НS КоА→ 
Реакция іс жүэінде қайтымсыэ рвакциялардың қатарына жатады. Ци-топлаэмаға коғарыда көрсетілген жолдармен түскен ацетил-КоА аце-тил-КоА-карбоксилаза ферментінің қатысуымен малонил-КоА-га ай-
налады:

Ацетил-КоА-қарбоксилаэа ферментінің коферменті биотиң, СО2 та-сымалдауға қатысады. Ацетил-КоА-карбоксилаэа ферменті реттеуші ферменттерге жатады. ферменттің активтілігін арттырушы актива-тордың ролін митохондрийдең цитоплазмаға келіп түсетін цитрат атқарады. Пальмитатсинтетаза фврменттін комплексі жеті ферменттен түрады және әрбір ферменттің атқаратын қызметі әртурлі Полифер-менттік комплекстің ортасында ацилді тасымаядаушы белок,ал жан-жагына қалған алты фермент орналасады. Схема түрінде пальмитат-синтетазалық ферменттік комплексті төмендегідей етіп бейнелеуге
болады
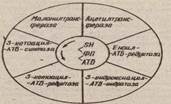
Ацил-тасымалдаушы белоқ /АТ6/ май кддщылы биосинтезінде синтеэ-дің аралық заттарын байланыетырып, оның әрі қарай журуіне мумкіндік береді, жалпы май қышқылы еинтеэінде орталық орын алады. АТБ-тың простетикалық тобының ролін 4-фосфопантотеин атқа-рады. Фосфопантотеиннің қурамында пантотеин қышқылы және ІН-тобы бар. АТФ-тың қурамындагы цистеин оны тагы да бір SН-тобымен қамтамасыэ етеді. Сондықтан ацил-тасымалдауда белок, әрі қарай -SН деп белгіленеді, Май қышқылы синтеэі темөндегідей био-химиялық реакцийлардың көмегімен журеді:
I/ Ацетил-КоА-ның қүрамындагы аңетил тобын пальмитсинтета-эа ферментімен байланыстыру:
 «
«
2/ Малонид-КоА-дан малонил іюбының синтетаза ферменттік комплексімен байланысуы,далонилтраңсацмлаза фермөнтінің қаты-суымен журеді

3) β-кетоацилсинтетаза ферментінің қатысуымен малонил мен ацетил топтарының С02 газын бөле конденсациялануы;

4/ β-кетоацил редуктаэа ферментінің жәнө НАдаН^Н^-тың қатысуымен тотықсыздану реакциясы:

5/ β-гидроксибутирил-АТВ қүрамындагы β-гидроксибутирил-гидроксиацилгидратаза фермөнтініңқатысуымен дегидратациялануы:

6/ НАДФН:Н+-тың көмегімен, тағы да тотықсыэдану реакциясы-
ның жүруі. •

Бутирил тобы ацетил-трансацилаэв ферментінің қатысуымен цистеиндік SН - тобына көшіріледі, пантотеиннің қурамындагы SН -келесі малонил тобын қосып алуға дайындалады.

Осымен бір цикл аяқталды. Реакция нәтижесінде, бутирилден боса-ған пантатеиннің SН- тобы келесі малонил тобын малонил- -КоА-дан қосып алады. Тізбек жоғарыда көрсетілген ферменттік реакциялардың көмегімен, тагы да екі көміртегі атомына ұзарады. Осындай 7 циклден кейін пальмитоил - -АТБ түзіледі. Пальмитоил --АТБ гидролиздік ферменттердің қатысуымең ацил-тасымалдаушы белоктан ыдырайды:

Түзілген пальмитин қышкшшнан әрі қарай көміртектік тіэбектің үзаруы нәтижесіңде басқа жоғары май қышқылдары түзілуі мүмкін.
5. Триглицеридтөр синтезі май т.б. тканьдерде май қоряарының жиналуы үшін қажет. Бүл синтез ішекке, май тканьдерінде, бүй-ректе, бауырда, бүлшықеттерде әртүрлі жолмен жүруі мүмкін. Триацилглицеридтер синтезі үшін глицернн жане стеарин, пальми-тин, олеин сияқты май қышқылдары қажет. Майлардың синтеэі гяи-церол-3-фосфаттыц синтезінен бастаяады. Бүйректе, ішек қабырға-ларында глицеролқиназа ферментінің активтілігі жоғары болғандық-тан майлардың синтеэі глицеринмен АТФ-тың арекеттесуінен баста-
лады.
Май тканьдерінде және бүлшықеттерде глицерол ферментінің активтілігі төмен болгандықтан, глицерол-3-фосфаггың түэіяуі гликолнэ және гликогенолиз ндтижесінде түэіяетін диоқсиацетон-фосфаттың НАД-қа тауелді глицерол-фосфатдегидрогеназаның қатысу-мен тотықсыздануы арқылы жүреді,
Ішекте сонымен қатар моноацетилглицеридтердвн диацилглице-ридтердің, триацилглицеридтердің түзілуі жүреді


6. Стериңдердің алмасуы. Холестерин ағзаға қажетті көптеген заттардың туындысы болып табылады (28-кесте).
Майлар мен липоидтер алмасуын реттеу орталығы гипоталамуста орналасқан. Майлар мен липоидтер алмасуын гипофиз, калқан, ұйкы және жыныс бездерінің гормондары реттейді.
Дата добавления: 2016-01-04; просмотров: 1; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!