Нуклеиновые кислоты. Системы репликации и репарации.
Глоссарий
Запаздывающая нить – цепь (нить) ДНК, которая синтезируется участками;
Лидирующая нить – цепь (нить) ДНК, которая синтезируется непрерывно во время репликации;
ДНК-полимераза (Polα) – разновидность фермента ДНК-полимераза, которая способная инициировать начало репликации;
ДНК-лигаза – фермент, катализирующий соединение двух молекул с образованием новой химической связи;
РНК-праймер – это короткий фрагмент нуклеиновой кислоты (олигонуклеотид), комплементарный ДНК- или РНК-мишени, служит затравкой для синтеза комплементарной цепи с помощью ДНК-полимеразы (при репликации ДНК);
Праймаза – фермент который синтезирует короткий фрагмент РНК, называемый праймером, комплементарный одноцепочечной матрице ДНК;
Фрагмент Оказаки – относительно короткие фрагменты ДНК (с РНК-праймером на 5' конце), которые образуются на отстающей цепи в процессе репликации ДНК;
ДНК-полимераза (Polδ) – разновидность фермента ДНК-полимераза, которая заполняет «пробелы» на этапе терминации.
Хеликаза – фермент, разрушающий фодородные связи между комплементарными цепями ДНК;
Топоизомераза – фермент, способный релаксироватьсверхспирализованные молекулы ДНК;
ДенатурацияДНК– расхождение цепей двухцепочечной молекулы ДНК вследствие экстремальных воздействий (температура, рН, денатурирующие агенты), что сопровождается потерей ее биологической активности.
|
|
|
ДНК
Это молекула, имеющая полимерную структуру. Мономером ДНК является нуклеотид. Каждый нуклеотид состоит из трёх компонентов: азотистое основание, дезоксирибоза и остаток фосфорной кислоты. Выделяют 4 типа нуклеотидов, чьи различия кроются в азотистых основаниях: аденин, тимин, цитозин и гуанин. Благодаря чередованию этих четырёх нуклеотидов и осуществляется кодирование наследственной информации.
ДНК представляет из себя двухцепочечную спираль. Каждая цепь содержит последовательность нуклеотидов, строго соответствующую последовательности другой цепи. Это соответствие достигается наличием водородных связей между направленными навстречу друг другу основаниями двух цепей: G и С или А и Т. Таким образом, цепи комплементарны (Рис. 3).
У каждой цепи различают 3’ и 5’ – концы.На Рис. 1. Можно отметить 5 концов, нумерация которых основана на нумерации атомов углерода в представленной молекуле. Связи между нуклеотидами в пределах одной цепи образуются путём соединения остатка фосфорной кислоты с 5’-концом углевода одного нуклеотида и 3’-концом углевода соседнего нуклеотида (Рис. 2). Такая химическая связь получила название фосфодиэфирной.
|
|
|
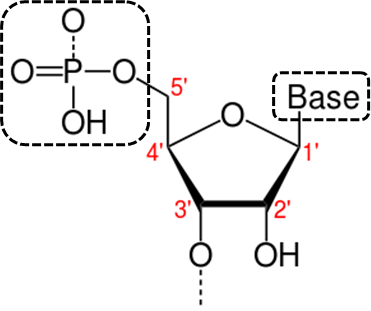 Рис. 1. Структура нуклеотида: левая рамка – остаток фосфорной кислоты, правая – азотистое основание (аденин, гуанин, цитозин, тимин, урацил); между ними – углевод
Рис. 1. Структура нуклеотида: левая рамка – остаток фосфорной кислоты, правая – азотистое основание (аденин, гуанин, цитозин, тимин, урацил); между ними – углевод
(https://ru.wikipedia.org/wiki/
Направленность).

Рис. 2. Схема связей нуклеотидов в одинарную цепь ДНК
(http://helpiks.org/4-955.html).
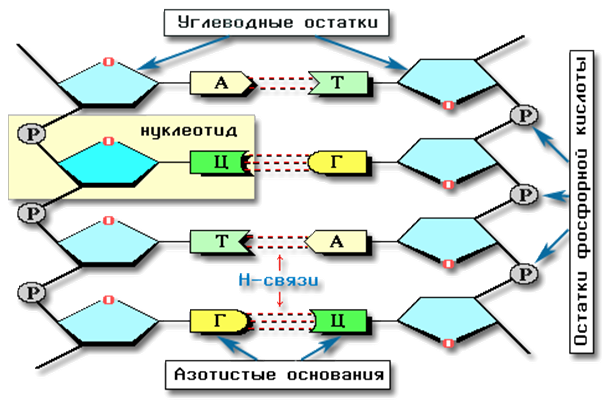
Рис. 3. Комплементарность в молекуле ДНК
(http://orgchem.ru/chem6/hm642.htm).
Репликация
Это процесс удвоения молекулы ДНК. Оно необходимо для последующего деления клетки. Если же не будет репликации, то с каждым новым клеточным циклом количество ДНК в клетке будет уменьшаться в два раза, чтобы этого избежать, и возник механизм удвоения молекулы.
Мы рассмотрим этот процесс в упрощённом виде, не углубляясь во все разновидности ферментов, участвующих в репликации.
Процесс можно разделить на 4 этапа:
1. Инициация;
2. Элонгация;
3. Исключение праймеров;
4. Терминация.
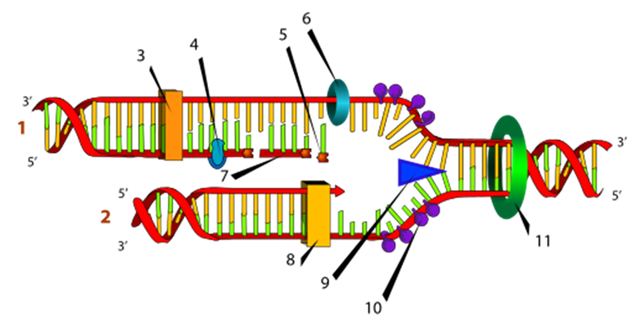
Рис. 4. Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздывающая нить, (2) лидирующая нить, (3) ДНК-полимераза (Polα), (4) ДНК-лигаза, (5) РНК-праймер, (6) праймаза, (7) фрагмент Оказаки, (8) ДНК-полимераза (Polδ), (9) хеликаза, (10) одиночная нить со связанными белками, (11) топоизомераза (http://nado.znate.ru/Репликация).
|
|
|
Инициация
Синтез новых одноцепочечных молекул ДНК может произойти только при расхождении родительских цепей. В определённом сайте (точка начала репликации) происходит локальная денатурация ДНК, цепи расходятся и образуются две репликативные вилки, движущиеся в противоположных направлениях.
Денатурация происходит под действием фермента хеликазы, который разрушает водородные связи между цепями, для этого он использует энергию АТФ.
Фермент топоизомеразарелаксируетсверхспирализованный участок молекулы, что необходимо для деятельности других ферментов. В результате происходит раскручивание участка суперспирализованной молекулы ДНК. В поддержании этого участка ДНК в раскрученном состоянии участвуют SSB-белки (от англ, singlestrandbindingproteins, т.е. белки, связывающиеся с одноцепочечными нитями ДНК). SSB-белки, не закрывая азотистых оснований, связываются с одноцепочечной ДНК по всей длине разделившихся цепей.
Элонгация
После того, как образованы достаточно свободные участки молекулы ДНК, к одноцепочечной ДНК присоединяется РНК-праймер. Дальнейшее формирование дочерней цепи будет происходить посредством последовательного присоединения нуклеотидов к 3’ – концу строящейся цепи.
|
|
|
На этом этапе важную роль играют ионы магния, они «активируют» нуклеотиды, нейтрализуя отрицательный заряд этих молекул, что увеличивает реакционную способность самих нуклеотидов.
Достраиванием дочерней цепи занимается фермент ДНК-полимераза, которая распознает азотистое основание матричной цепи и строит напротив него нуклеотид комплементарный, распознанному азотистому основанию.
Исключение праймеров

Рис. 5. Процесс исключения РНК-праймеров из дочерних цепей.
Короткие РНК-фрагменты имеют иную, чем у ДНК структуру, поэтому не являются её основной составляющей. Поэтому после того, как «застроены» участки между РНК-праймерами, последние исключаются из дочерних цепей. На их месте возникают «пробелы».
Терминация
Естественно, в ходе молекулы ДНК не могут присутствовать промежутки между нуклеотидами, поэтому такие «разрывы» застраиваются ДНК-полимеразой. На данном этапе праймерами выступают уже построенные участки дочерней цепи ДНК. Однако, как видно из рисунка 3 на концах дочерних цепей остаются недостроенные участки.
Это не является аномалией. Такое укорачивание происходит на участках, в которых не содержатся кодирующие признаки участки ДНК. Укорачивание идёт за счёт повторяющихся последовательностей. Это своего рода запас нуклеотидов, которые ДНК неизбежно теряет с каждым делением, но эта утрата не сказывается на кодировании признаков. Эти участки называются теломерами. Существуют механизмы восстановления теломерных участков, этим занимается фермент теломераза, но большинство соматических клеток лишены активности этого фермента. Одной из функцией теломер называют регуляцию клеточного старения, это объясняется тем, что когда теломеры достигают своих минимальных размеров, то клетка перестаёт делиться.
Репарация
Это механизм восстановления структуры и/или функционала повреждённой молекулы ДНК. Нарушение работы этого механизма может вести к развитию различного рода заболеваний.
Одним из примеров экстренных исправлений является SOS–репарация. Её значение проявляется в момент, когда клетка стоит на пороге репликации. Если к этому моменту имеются ошибки в молекуле ДНК, то это может привести к остановке процесса удвоения нуклеиновой кислоты. В случае такой остановки нормальное деление клетки и последующая жизнедеятельность невозможны. Таким образом эта система обеспечивает выживание клетки на этапе репликации, даже при наличии ошибок в ДНК.
Суть механизма проста. При активации SOS-системы начинается синтез белков, которые соединяются с ДНК-полимеразным комплексом и делают его «невнимательным» к имеющимся ошибкам. ДНК-полимеразный комплекс, в свою очередь, строит дочернюю цепь напротив дефектных участков матричной цепи.
Однако, есть ряд нюансов. Во-первых, такой способ чреват сохранением ошибок ДНК. Во-вторых, если эти ошибки критичны, то после деления клетка всё равно погибнет. В-третьих, даже если клетка не гибнет после деления, сохраняется риск утраты клеткой ряда своих функций.
РНК
Является разновидностью нуклеиновых кислот. Отличительными особенностями (от ДНК) является одноцепочечная структура, вместо дезоксирибозы – рибоза, также отличается одно азотистое основание (вместо тимина – урацил). Общей функцией всех видов РНК можно назвать реализацию генетической информации в признаки.
Таблица 1. Виды РНК и их функции.
| Виды: | Функции |
| гетероядерная (гяРНК), | переносят генетическую информацию от ядерной ДНК к цитоплазме. |
| информационная (иРНК или мРНК) | Информационная РНК образуется в результате процессинга гяРНК, при котором происходят вырезание некодирующих участков (интронов) и склеивание кодирующих экзонов. Таким образом, в состав иРНК входят кодирующая информация соответствующих видов гяРНК, а также фланкирующий лидерный и трейлерный участки, по этой причине она значительно короче. |
| транспортная (тРНК), | «заряженная» молекула несёт на своём 3'-конце аминокислоту, а посередине конформационной структуры «клеверный лист» находятся три специфических основания, именуемые антикодоном. Последовательность оснований в антикодоне напрямую зависит от вида аминокислоты, прикреплённой к 3'-концу. |
| рибосомная (рРНК), | Каждая рибосома состоит из большой и малой субъединиц. Они содержат большое количество белков, синтезированных посредством трансляции иРНК, а также РНК, которая не подвергается трансляции. Термин «рибосомальная РНК» относят именно к нетранслируемому материалу. |
| малая ядерная РНК (snPHK), | Превращение гяРНК в иРНК путём удаления интронов проходит в ядерном комплексе РНК-белков, называемом сплайсомой. У каждой сплайсомы есть ядро, состоящее из трёх малых (низкомолекулярных) ядерных рибонуклео-протеинов, или снурпов. Каждый снурп содержит хотя бы одну малую ядерную РНК и несколько белков. Основная функция — распознавание специфических рибонуклеиновых последовательностей посредством спаривания оснований по типу РНК—РНК. |
| малая ядрышковая РНК (snoPHK), | Участвует в направлении или проведении модификаций оснований в рРНК и малой ядерной РНК, таких, как, например, метилирование и псевдоуридинизация. |
| Сигналраспоз- нающая РНК (srpPHK), | Распознаёт сигнальную последовательность белков, предназначенных для экспрессии, и участвует в их переносе через цитоплазматическую мембрану. |
| микро-РНК (miPHK), | Контролируют трансляцию структурных генов путём комплементарного связывания с З'-концами нетранслируемых участков иРНК. |
Транскрипция
Механизм построения молекулы РНК с использованием молекулы ДНК, как образца. В ряде источников это называют переносом генетической информации с ДНК на РНК.
Процесс транскрипции аналогичен репликации. Также выделяют инициацию, элонгацию и терминацию. Ключевыми отличиями является то, что вместо дочерней ДНК синтезируется РНК, также отличается основной действующий фермент (вместо ДНК-полимеразы работает РНК-полимераза).
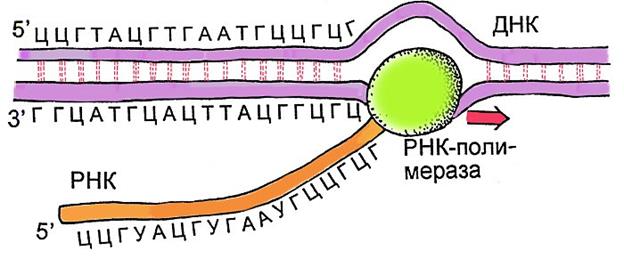
Рис. 6. Схема транскрипции
(http://pisum.bionet.nsc.ru/kosterin/lectures/lecture4/lecture4.htm).
Трансляция
Это процесс синтеза полипептидной цепи в рибосоме. В этом механизме принимают участие три основных вида РНК (информационная, рибосомальная и транспортная. иРНК приносит в рибосому информацию об аминокислотной последовательности. рРНК входит в состав рибосомы, не кодирует информацию, участвует в процессе образования полипептидной цепи (возникновении последовательных связей между аминокислотами). Транспортная РНК доставляет аминокислоты в рибосому.
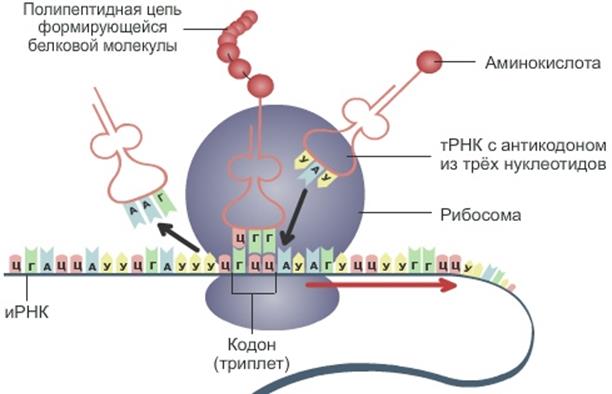
Рис. 7. Схема процесса трансляции.
Вопросы для самоподготовки:
1. Строение нуклеотида?
2. Система репарации?
3. Этапы репликации?
4. Виды и функции РНК?
Дата добавления: 2022-11-11; просмотров: 32; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!