Факторы, влияющие на эффективность обора
Лекция – 9
Редакция 03.03.2022 г.
Тема 2: Современные методы оценки эффективности отбора
Вопросы
1. Теоретические основы вычисления оценок эффективности отбора.
2. Расчет величины селекционного дифференциала.
3. Факторы, влияющие на эффективность отбора.
Ключевые слова:
отбор, эффект селекции, селекционный дифференциал, интенсивность отбора, измерение и прогноз действия отбора, повышение селекционного дифференциала.
Распределение бюджета рабочего времени
Форма проведения занятий – аудиторные занятия, лекция.
Бюджет времени – 2 часа.
Количество двухчасовых занятий – 1.
Распределение бюджета времени:
- 1 час: рассмотрение ключевых понятий и теоретических основ формирования оценок эффективности отбора в лесной селекции;
- 1 час: рассмотрение алгоритмов вычисления оценок эффективности отбора в лесной селекции.
Литература
1. Гужов Ю.Л., Фукс А., Валичек П. Селекция и семеноводство культурных растений. – М.: Агропромиздат,1991. – 463 с. – (Учебники и учеб. Пособия для студентов высш. Учеб. Заведений).
2. Бессчетнов В.П., Кентбаев Е.Ж., Бессчетнова Н.Н, Кентбаева Б.А. Лесная селекция / В.П. Бессчетнов, Е.Ж. Кентбаев, Н.Н Бессчетнова, Б.А Кентбаева. - Учебник для студентов лесохозяйственных специальностей. Гриф Республиканского учебно-методического совета Республики Казахстан. – Алматы: Нур-Принт , 2015. – 358 с.
|
|
|
3. Бессчетнова Н.Н. Сосна обыкновенная (Pinus sylvestris L.). Селекционный потенциал плюсовых деревьев / Н.Н. Бессчетнова. – Saarbrücken: Lap Lambert Academic Publishing GmbH & co. KG. ISBN 978-3-8443-5608-3, 2011 – 402 c.
4. Бессчетнова Н.Н., В.П. Бессчетнов. Сосна обыкновенная (Pinus sylvestris L.). Морфометрия и физиология хвои плюсовых деревьев / Н.Н. Бессчетнова, В.П. Бессчетнов. Монография. – Нижний Новгород: Нижегородская государственная сельскохозяйственная академия, 2014. – 368 с.
5. Бессчетнова Н.Н. Сосна обыкновенная (Pinus sylvestris L.). Репродуктивный потенциал плюсовых деревьев / Н.Н. Бессчетнова. Монография. – Нижний Новгород: Нижегородская государственная сельскохозяйственная академия, 2015. – 586 с.
6. Бессчетнова Н.Н. Сосна обыкновенная (Pinus sylvestris L.). Эффективность отбора плюсовых деревьев / Н.Н. Бессчетнова. Монография. – Нижний Новгород: Нижегородская государственная сельскохозяйственная академия, 2016. – 382 с.
7. Бессчетнов В.П. Полиморфизм казахстанских популяций облепихи крушиновидной (Hippophae rhamnoides L.) по хозяйственным и адаптивным признакам. Дис. док. биол. наук. Алматы, 1994. – 543 с.
8. Бессчетнов В.П. Полиморфизм казахстанских популяций облепихи крушиновидной (Hippophae rhamnoides L.) по хозяйственным и адаптивным признакам. Дис. док. биол. наук. Алматы, 1994. – 543 с.
|
|
|
9. Бессчетнов В.П.
10. Чмыр А.Ф., Бессчетнов В.П. Экология и культура облепихи Hippophae rhamnoides L. – СПб, СПбНИИЛХ, 1998. – 278 с.
11. Котов М.М. Генетика и селекция. Часть 1.: Учебник для вузов. – Йошкар-Ола: МарГТУ, 1997. – 280 с.
12. Котов М.М. Генетика и селекция. Часть 2.: Учебник для вузов. – Йошкар-Ола: МарГТУ, 1997. – 108 с.
13. Любавская А.Я. Лесная селекция и генетика.: Учебник для вузов. – М.: Лесная промышленность, 1982. – 288 с.
14. Джонатан В. Райт. Введение в лесную генетику. М.: Лесная промышленность, 1978. – 470 с.
Асинхронно-синхронная связь с преподавателем:
1. Бессчетнов Владимир Петрович: E-mail: lesfak@bk.ru
Мобильный телефон: 8-906-355-81-97.
2. Бессчетнова Наталья Николаевна: E-mail: besschetnova1966@mail.ru
Мобильный телефон: 8-960-160-54-32.
Теоретические основы вычисления оценок эффективности
Отбора
Оценка эффективности отбора представляет собой одну из важных задач селекции, в частности, селекции методом отбора (а в широком смысле – селекции как формы деятельности человека, в целом). Для её реализации используют специальные оценочные критерии, называемые селекционным дифференциалом и интенсивностью отбора. Проведение отбора по фенотипу, в частности, отбор плюсовых деревьев, потомство которых используется для создания лесосеменных плантаций первого порядка, будет эффективным при высоком уровне аддитивной (не связанной с эффектом доминирования, а определяемой собственно набором генов) наследственности, а также будет зависеть от проявления эффектов межаллельного взаимодействия, в целом: доминирования (полного или неполного), кодоминирования, сверхдоминирования. В самом общемсмысле этот фактор влияния на результаты отбора рассматривают как генотипический. Аддитивная наследственность в селекции рассматривается как наследственность, обусловленная собственно набором генов, без учета эффекта их аллельного (прежде всего, доминирование) взаимодействия. Такая наследственность часто называется истинной и оценивается коэффициентом наследуемости в узком смысле. Аддитивная наследственность – это наследственность, обусловленная суммирующим эффектом действия полимерных генов (расширить и уточнить определение понятия аддитивная наследственность и понятия неаддитивная наследственность; возможно эпистаз, плейотропия, полимерная наследственность, комплементарность, гены модификаторы!!!).
|
|
|
Кроме того, эффективность отбора будет определяться тем, насколько велико влияние факторов среды на формирование фенотипических признаков, т.е. будет зависеть от так называемой средовой компоненты вариансы или остаточной дисперсии признака. Влияние среды выражается в возникновении (приобретении, формировании, проявлении…) у представителей популяции разнообразных ненаследуемых изменений (уклонений от среднего…) значения признака. В целом, они (флуктуации) расширяют фенотипическое разнообразие исходного материала. Однако целью и желаемым результатом селекции всегда является отбор лучших генотипов, а не фенотипов (о чем мы уже говорили, рассматривая проблемы, стоящие перед современной селекцией). Модифицирующее влияние среды в такой ситуации будет усложнять работу по отбору лучших генотипов. Чем сильнее выражено такое модифицирующее влияние среды, тем сложнее организовать эффективный отбор лучших генотипов, тем больше шансов при отборе по фенотипу, что имеет место в лесной селекции, «отобрать» не собственно «лучший» генотип, а лишь видимую нами ненаследуемую часть фенотипических проявлений признака, которая не сохранится при семенном размножении или при изменении условий среды (в иных условиях...) Это в конечном итоге снизит результаты или эффективность отбора, если под отбором понимать именно отбор лучших генотипов.
|
|
|
Напряженность или уровень интенсивности отбора зависит от того, какая часть (10 %, 5 % или 1 %) или, сколько наилучших особей будет отобрано из популяции (из исходной совокупности в общем случае), например: 15 самых лучших или 10 самых лучших, или только 5 самых лучших, или в конечном итоге только одно (1) самое лучшее дерево. Уровень интенсивности отбора зависит и от того, какой предел проявления признака признается достаточным для включения особи в число отобранных по лучшим характеристикам: все особи с высотой 32 м и более, или все особи с высотой 33 м и более, или, наконец, все особи с высотой 35 м и более и т.п.
Результативность отбора обусловлена и его устанавливаемым уровнем (заданной «жесткостью» или «строгостью» или «интенсивностью» отбора), который в свою очередь определяется задачами селекции и принятым в каждом конкретном случае (сосна обыкновенная в лесотундре или в Брянской области) порядком проведения селекционных мероприятий. В конечном итоге, определяется селекционером. Так, селекционный дифференциал при его заданном уровне используется при выделении плюсовых деревьев. Действующими нормативами (Указания…, 2000; Приказ…, 2015) (дать расширенную трактовку «Правил» и заложенных в них нормативов!!!) устанавливается величина селекционного дифференциала в процентах, который должен быть достигнут при отборе плюсового дерева. По высоте оно должно превосходить среднее значение в насаждении на 10 %, а по диаметру – на 30 %. Ранее существовали еще более жесткие требования к отбираемым плюсовым деревьям (Вересин, 1963; Временные рекомендации…, 1966; Указания о порядке отбора…, 1972; Основные положения по лесному семеноводству…, 1976; 1991): по высоте 20 – 25%, по диаметру 30 – 35%. Эти определяемые нормативом превышения значений установленных признаков (высоты и диаметра) и представляют собой селекционный дифференциал, заданный при отборе плюсовых деревьев.
Для аллогамных популяций древесных и кустарниковых пород характерно промежуточное наследование, когда рецессивный ген не в полной мере блокируется доминантным, и полигенная аддитивная наследственность большинства количественных признаков, что определено эволюционным процессом. Полимерный аддитивный (уточнить корректность терминов!!!) характер наследования основных количественных признаков деревьев и кустарников порождает клинальную изменчивость и приводит к возникновению множества постепенно переходящих друг в друга его количественных проявлений (чем больше полимерно действующих генов находится в доминантном аллельном состоянии, тем сильнее проявление полигенно наследуемого признака).
При отборе по фенотипу (отбор плюсовых деревьев) результат будет определяться рядом условий:
- тем, насколько велика доля наследственной обусловленности проявлений признака или, что то же самое, генотипической обусловленности изменчивости признака, в общей, учитываемой нами при отборе фенотипической изменчивости признака у селектируемых особей, от степени генетической детерминированности признака, от степени его экотипической стабильности;
- тем, какова доля средовой дисперсии признака или доля влияния факторов среды на его проявление, от его лабильности, от подверженности признака влиянию факторов среды.
В конечном итоге фенотипическое проявление признака, то, на что направлен отбор по фенотипу (в лесной селекции – это отбор плюсовых деревьев), будет зависеть от соотношения долей влияния на проявление фенотипа факторов наследственности (генотипа в широком смысле) и факторов среды (доля ненаследственной изменчивости). Это позволяет рассматривать взаимное действие (оно всегда именно таково) факторов среды и факторов наследственности на формирование фенотипа особи как действие одного фактора – соотношения между долями влияния среды и генотипа. Тогда эффект отбора по количественным признакам можно представить как результат, зависящий от действия двух формально независимых друг от друга комплексных факторов:
- от интенсивности отбора, определяемой задачами селекции и планом селекционных мероприятий;
- от силы модифицирующего влияния среды, под которой понимают долю ненаследственной изменчивости в общей фенотипической изменчивости, рассматривая её как оценку соотношения «наследственность/среда».
Доля влияния фактора среды на формирование общей фенотипической дисперсии, формально зависит от величины генотипической вариансы и наоборот (поскольку они являются долевыми оценками). Относительную силу генетического влияния и влияния внешней среды на фенотипическое проявление признака оценивают с помощью коэффициента наследуемости (H2), который выражается в долях от единицы или в процентах. Например, Н2 = 0,78 или 78%. Это значит, что проявление признака на 78% обусловлено генотипом особи, а на 22% – условиями среды. Для планирования процесса отбора и прогнозирования его результатов важно дать оценку влияния этих двух факторов. Эффективность или результативность отбора можно оценить двумя широко применяемыми показателями:
- селекционным дифференциалом (обозначим его S);
- интенсивностью отбора (обозначим его i).
В большинстве отечественных публикаций эти два показателя оценок эффективности отбора рассматриваются как самостоятельные величины, а в ряде зарубежных публикаций (Райт, 1975) – как одна величина, выраженная в разной форме. Согласно устоявшимся представлениям, селекционный дифференциал – это разность между средней величиной признака в группе отобранных особей (плюсовых деревьев, семей, клонов) и соответствующей средней величиной того же признака в исходной популяции (совокупности). Его вычисляют по весьма простой формуле: S = 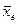 -
- 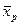 ,
,  где:
где:
- 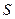 – селекционный дифференциал;
– селекционный дифференциал;
- – среднее значение признака в группе отобранных растений (индекс е от extraction – выделение, экстракция) или значение одного растения, которыми могут быть отобранные по фенотипу плюсовые деревья, отобранные лучшие семь в испытании семенных потомств, отобранные лучшие вегетативные потомства в клоновом отборе;
- – среднее значение признака в популяции, из которой отобрана группа особей (индекс р от population – популяция), рассматривается как собственно природная популяция, так и как любая исходная группа особей, из числа которой производят отбор.
Для уточнения статистической значимости оценок селекционного дифференциала применяют критерий Стьютента, вычисление которого ведут по формуле достоверности разности двух средних:
 ,
,
где:
- t – расчетное значение критерия Стьютента
- - – разность средних значение признака в группе отобранных особей и в исходной популяции (совокупности), при этом - = и представляет собой селекционный дифференциал;
- – среднее значение признака в группе отобранных растений (индекс е от extraction – выделение, экстракция) или значение одного растения, которыми могут быть отобранные по фенотипу плюсовые деревья, отобранные лучшие семь в испытании семенных потомств, отобранные лучшие вегетативные потомства в клоновом отборе;
- – среднее значение признака в исходной популяции, из которой отобрана группа особей (индекс р от population – популяция), рассматривается как собственно природная популяция, так и как любая исходная группа особей, из числа которой производят отбор.
- m 2 e – возведенная в квадрат абсолютная ошибка среднего значения признака в группе отобранных особей;
- m 2 p – возведенная в квадрат абсолютная ошибка среднего значения признака в исходной популяции, из которой отобрана группа особей;
- Ne – численность группы отобранных особей;
- Ne – численность особей в исходной популяции (совокупности);
- ν – число степеней свободы для определения табличных значений критерия Стьюдента: ν = Ne + Np – 2 = (Ne – 1) + (Np – 1).
Если вычисленное значение t-критерия больше его табличного значения для определенного числа степеней свободы и заданного уровня точности, то различия между средними можно признать достоверными. Такая ситуация в смысловом отношении означает то, что отбор действительно привел к изменению средних значений селектируемого признака в отобранной группе по отношению к исходной совокупности, то, что величина полученного селекционного материала достоверно превышает среднее популяционное значение того же признака в исходной популяции, из которой был осуществлен отбор. Это соответствует тому, что разница между средними превышает величину случайных отклонений, не связанных с действием рассматриваемого фактора (в нашем случае с действием отбора). Это является подтверждением результативности отбора. Данный показатель можно рассматривать как критерий достоверности селекционного дифференциала.
Традиционно селекционный дифференциал применяют для оценки эффективности отбора, в том числе, отбора по фенотипу плюсовых деревьев, отбора, проводимого по результатам испытания потомств – семей, или клонов. Например, если средняя высота деревьев в насаждении (популяции) составляет 30 метров, а средняя высота группы отобранных особей – 33 метра, то селекционный дифференциал будет равен: 33 – 30 = 3 м, или 10% (от исходной популяционной величины). Если при испытании семенного потомства (испытательные культуры сосны обыкновенной в Семеновском лесничестве Нижегородской области) средняя высота всего семенного потомства в испытательных культурах 20 м, а высота сопоставляемого с ним потомства от одного плюсового дерева (например, плюсового дерева № 19) – 21 м, то селекционный дифференциал достигнет: 21 – 20 = 1 м, или 5%.
Выражают селекционный дифференциал в натуральных единицах измерения значений признака, например: высота – в м; диаметр ствола на высоте 1.3 метра – в см; объем ствола – в м3; площадь поперечного сечения ствола – в см2 и т.д. При этом могут также использоваться как относительные размерные показатели (отношение высоты ствола, выраженной в метрах к площади поперечного сечения ствола, выраженной в см2 – м/см2; плотность древесины – г/см3 и т.д.), так и безразмерные величины (отношение длины листовой пластинки, выраженной в см, к ширине листовой пластинки, выраженной в сантиметрах, называется коэффициентом формы и выражается безразмерной величиной; аналогично выражается коэффициент формы шишек или семян и т.п.). Эта форма выражения селекционного дифференциала наиболее предметна, наглядна и легко воспринимается. Она позволяет оценить эффективность отбора, проводимого на разных объектах, но по одному и тому же показателю, например, по высоте ствола и его объему. Вместе с тем понятно, что такая форма выражения селекционного дифференциала не позволяет сопоставлять эффективность отбора, проводимого по разным признакам: высоте ствола (в метрах) и диаметру ствола (в сантиметрах), даже если отбор ведется в одной и той же популяции или насаждении.
В этой связи для получения сопоставимой оценки эффективности отбора, проводимого по разным признакам, селекционный дифференциал выражают в процентах от среднего значения признака в исходной совокупности – в популяции, из которой проведен отбор. Понятно, что при такой оценке в относительных величинах (процент от числа есть величина относительная или доля) дать сравнительную оценку эффективности отбора любых самых разных признаков, и указать, по какому признаку отбор оказался более эффективным, а по какому – менее эффективным, достаточно просто. При подтверждении достоверности селекционного дифференциала производят соответствующие вычисления его значения в относительных величинах: в % от среднего значения признака в исходной популяции. Используем формулу: S% = 100×( - )/ , где:
- S % – селекционный дифференциал, выраженный в % от среднего популяционного значения;
- - – разность средних значение признака в группе отобранных особей и в исходной популяции (совокупности), при этом - = и представляет собой селекционный дифференциал;
- – среднее значение признака в группе отобранных растений (индекс е от extraction – выделение, экстракция) или значение одного растения, которыми могут быть отобранные по фенотипу плюсовые деревья, отобранные лучшие семь в испытании семенных потомств, отобранные лучшие вегетативные потомства в клоновом отборе;
- – среднее значение признака в исходной популяции, из которой отобрана группа особей (индекс р от population – популяция), рассматривается как собственно природная популяция, так и как любая исходная группа особей, из числа которой производят отбор.
Селекционный дифференциал, выраженный в натуральных величинах, как мера интенсивности отбора имеет серьезный недостаток: по величине селекционного дифференциала результативность (строгость) отбора по определенному признаку в различных популяциях можно сравнить только в том случае, если изучаемые популяции имеют сопоставимую или одинаковую изменчивость определенного признака. В природе данное условие обычно не обнаруживается, и популяции, как правило, характеризуются различными уровнями изменчивости по анализируемым признакам. Это справедливо в отношении практически всех хозяйственно важных и адаптивных признаков у большинства лесных деревьев и кустарников. Однако, хорошо известно то, что оценка эффективности отбора зависит от дисперсии признака. Понятно, что по признаку с большей дисперсией будет достигнут и заведомо больший результат, и, наоборот – для признака с меньшей дисперсией результат окажется меньшим. Тогда сравнение эффективности отбора по признакам, имеющим разную дисперсию, будет не вполне корректным, даже если её оценку выразить в процентах.
Для преодоления этого обстоятельства при получении сопоставимых оценок эффективности отбора, независимых от уровня изменчивости признака, применяют показатель, выражающий селекционный дифференциал в долях от среднего квадратического отклонения данного признака в исходной популяции (мы обозначили его как s р). Этот показатель получил название «интенсивность отбора». Интенсивность отбора – это показатель, определяющий, какую долю от среднеквадратического отклонения признака в исходной популяции составляет достигнутый при отборе селекционный дифференциал. Он представляет собой отношение величины селекционного дифференциала, выраженного в натуральных единицах измерения, к величине среднеквадратического отклонения (как оценке дисперсии…) признака в исходной популяции, из которой был произведен отбор, выраженной в тех же единицах измерения. Формула его расчета достаточна проста: i = S/s р, где:
- i – интенсивность отбора;
- S – селекционный дифференциал;
- σp – среднее квадратическое отклонение признака в популяции.
Интенсивность отбора показывает, какое число «сигм» содержит селекционный дифференциал. Легко заметить, что она является безразмерной величиной, поскольку в получаемом отношении участвуют только одноразмерные величины, при этом не зависящая от дисперсии (уровня изменчивости…) признака в разных популяциях. Такие свойства рассматриваемого показателя позволяют сравнивать интенсивность отбора в любом числе популяций при специфике характера изменчивости признака в каждой из них.
Факторы, влияющие на эффективность обора
На эффективность отбора оказывают влияние как особенности строения (организации…) популяций (их структура или характер распределения частот, дисперсия признаков особей), так и заданный уровень интенсивности отбора.
Во многом результативность отбора связана с наличием достаточного исходного материала для селекции. Если отбор осуществляется из широкой базы исходного материала – из большого числа особей (сеянцы в питомнике, деревья в насаждении) или из многочисленной популяции, то его эффективность будет выше. Действительно, с большей вероятностью отобрать что-либо с высоким значением признака можно из большого числа единиц отбора. Наиболее выдающиеся по значению своих признаков особи встречаются редко, и шанс их найти тем больше (выше…), чем больше растений будет подвергнуто отбору.
Селекционный дифференциал можно рассматривать как функцию числа отобранных деревьев (или семенных семей, или клонов) и числа деревьев (или семей, или клонов), из которых провели отбор. При большем отношении числа изученных деревьев (числа деревьев в популяции, которая подверглась отбору) к отобранным деревьям селекционный дифференциал выше. Однако связь между ними не прямая. Джонатан В. Райт (1978, стр. 161) в своей работе приводит таблицу взаимоотношения между селекционным дифференциалом и числом деревьев в популяции, которое следует подвергнуть обмерам при отборе (табл. 1): из которого следует осуществить отбор с максимальной жесткостью (отобрать одно самое лучшее дерево), чтобы достичь установленного уровня эффективности отбора, выраженного в «сигмах».
Для достижения эффективности отбора, при котором селекционный дифференциал (изменение величин среднего значения признака вследствие отбора) достигнет уровня одного среднеквадратического отклонения, численность исходной совокупности, которая подвергается отбору, должна составить 4 будет отобрано 1 лучшее из 4.
Таблица 1
Связь между селекционным дифференциалом (в долях от стандартного отклонения) и числом деревьев, которые надо изучить (учесть) при отборе
| Селекционный дифференциал | Число деревьев | Селекционный дифференциал | Число деревьев | Селекционный дифференциал | Число деревьев |
| 1,0 | 4 | 2,5 | 159 | 4,0 | 31540 |
| 1,5 | 13 | 3,0 | 739 | 5,0 | 3588000 |
| 2,0 | 42 | 3,5 | 4298 | 6,0 | 100000000 |
Легко заметить, что число биометрированных деревьев (отношение…) следует увеличить приблизительно в 10 раз, для того чтобы селекционный дифференциал стал равен 2 σ, а не 1 σ; для того, чтобы селекционный дифференциал увеличить с 2 до 3 величин стандартных отклонений число деревьев в исходной совокупности должно быть увеличено в 17,6 раза, а для того, чтобы селекционный дифференциал возрос с 3 до 4 стандартных отклонений число деревьев необходимо увеличить в 42,7 раза. Следует иметь в виду, что приведенные в таблице 1 величины достоверны только в том случае, если отбор проводят в сходных условиях. Как видим, на практике может возникнуть ограничение селекционного дифференциала, вызванное большой трудоемкостью работ и обширными площадями обследований. Тем не менее, практически всегда можно сравнить деревья, растущие в нескольких десятках метров одно от другого, и предположить (с той или иной степенью вероятности), что выбрано самое лучшее из них. Ограничение величины селекционного дифференциала целесообразно и в целях экономии: слишком дорого обходится обследование сотен или тысяч гектаров для отбора одного дерева.
При массовом отборе в питомнике или при отборе растений, устойчивых к болезням в условиях эпидемии, селекционный дифференциал равен 3 – 4 σ; при массовом отборе в однородном коренном лесу или культурах общего назначения, созданных сеянцами из нормальных семян, (селекционно не улучшенных…), селекционный дифференциал составляет 2 – 3 σ; при семейственном отборе или отборе для испытания потомства внутри семьи селекционный дифференциал равен 1 – 2 σ.
Значение селекционного дифференциала как показателя, применяемого на практике, можно оценить на следующем примере (по Райту, 1978). Пусть в испытательных культурах, выращенных в повторностях с соблюдением всех требований, представлены семенные потомства 10 плюсовых деревьев сосны обыкновенной из левобережья р. Волги (Семеновский специализированный семеноводческий лесхоз). Обозначим условно эти плюсовые деревья буквами английского алфавита от А до J. Пусть оценка относительной высоты отобранных деревьев будет представлена в таблице следующим образом (табл. 2).
Таблица 2.
Дата добавления: 2022-11-11; просмотров: 112; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!