ХАРАКТЕРИСТИКИ ВЕЛИЧИНЫ И СТРУКТУРЫ ПОПУЛЯЦИИ
ЛЕКЦИЯ 5-8
Экология популяций
ПЛАН:
1) Определение популяции
2) Характеристики величины и структуры популяции
2.1 Величина популяции
2.2 Пространственное распределение особей в популяции
2.3 Плотность популяции
2.4 Возрастная структура популяции
2.5 Половая структура популяций
2.6 Этологическая структура популяций
2.7Генетическая структура популяций
3. Динамика популяции
3.1 Рождаемость и смертность. Кривые выживания
3.2 Миграции
3.3 Рост численности популяции
3.4 r, K и Lстратегии
3.5 Внутрипопуляционные взаимодействия
3.6 Колебания численности популяции
4.Принципы управления популяцией
4.1 Популяция как единица эксплуатации
4.2 Популяция как единица регулирования численности
4.3 Популяция как единица охраны.
ОПРЕДЕЛЕНИЕ ПОПУЛЯЦИИ
Термин «популяция» (от лат. populus – народ, население) появился в конце ХVIII в. и долгое время использовался достаточно произвольно: для обозначения любой группы особей, которой оперировал исследователь, либо совокупности особей, населяющих определенное пространство. В тот же период в англоязычной литературе в схожем контексте нередко применялся английский термин population – как эквивалент русским словам «население» или «народонаселение» (например, в очень известном трактате Т. Мальтуса «Эссе о принципах народонаселения» (Essay on the principle of population, 1798). Однако постепенно к середине ХХ в. складывалось более узкое понимание термина «популяция», причем в разных областях биологической науки оно заметно отличалось. Так, в популяционной генетике под популяцией понимали систему генотипов, посредством свободных скрещиваний связанных в единое целое, в экологии – любую группу организмов одного вида, занимающую определенное пространство и функционирующую как часть биотического сообщества. Для того чтобы подчеркнуть экологический контекст понимания этого термина, стали использовать словосочетания «экологическая популяция» и «элементарная популяция». Отметим, что в ХХ в. как в экологии, так и в генетике понятие популяции подразумевало именно совокупность особей одного вида.
|
|
|
В середине ХХ в. термин «популяция» стал одним из ключевых в теории эволюции. Именно в этот период благодаря работам Н. В. Тимофеева-Ресовского и ряда других авторов оформился взгляд на популяцию как элементарную эволюционную единицу, способную реагировать на изменение среды перестройкой своего генофонда. Благодаря этому возникло и стало быстро развиваться самостоятельное комплексное направление в биологической науке – популяционная биология, объединившая популяционную экологию, генетику популяций и учение о микроэволюции, наряду с которыми сформировался целый ряд новых научных дисциплин и подходов (эволюционная экология, популяционная морфология, биохорология, фенетика и др.).
|
|
|
По этой причине современное определение популяции должно исходить с позиций популяционной биологии. Такому требованию в наибольшей степени соответствует определение, данное в 1987 г. А. В. Яблоковым: популяция – минимальная самовоспроизводящаяся группа особей одного вида, на протяжении эволюционно длительного времени населяющая определенное пространство, образующая самостоятельную генетическую систему и формирующая собственное экологическое гиперпространство.
Как видно, от приведенного выше узкоэкологического понимания популяции современная ее трактовка отличается рядом условий:
1) популяция представляет собой самостоятельную генетическую систему и характеризуется единым и в определенной степени своеобразным генофондом, который может выражаться как в особенностях соотношения разных генотипов, так и в наличии тех или иных специфических генов-маркеров;
2) для популяции в определенной степени необходимо быть изолированной от других популяций этого же биологического вида, причем степень изоляции должна быть достаточной для устойчивого сохранения генофонда популяции в ее географических границах;
|
|
|
3) популяция со всеми ее отличительными особенностями сохраняется на протяжении большого (эволюционно значимого) числа поколений.
Принимая во внимание эти условия, мы не можем считать самостоятельной популяцией произвольно взятую группу особей какого-то вида, существующую относительно короткое время даже при достаточно выраженной пространственной изоляции (например, жужелиц одного вида, населяющих газон в центре города). Нет достаточных оснований считать самостоятельной популяцией и группировку особей, не обладающую устойчивым генетическим своеобразием, даже если она характеризуется очевидными экологическими особенностями (пример – группа сосен, произрастающих на небольшом верховом болоте, окруженном сухими сосновыми борами, – если только не будет установлен факт устойчивой генетической изолированности их от окружающих деревьев того же вида).
Впрочем, пока нельзя утверждать, что очерченная данным определением концепция популяции уже стала общепринятой. Так, ученые-экологи в рамках конкретных исследований зачастую исходят из более упрощенного восприятия популяции как некой группы особей одного вида, существующей и функционирующей в рамках конкретного сообщества, абстрагируясь при этом от таких характеристик, как совокупная величина популяции, наличие или отсутствие реальных изолирующих барьеров между ее частями, своеобразие генофонда и т. п. Такой подход вполне допустим, но требует терминологических оговорок. По этой причине в научном обиходе в последние десятилетия стали использоваться такие понятия, как географическая популяция (группа особей одного вида, заселяющая территорию с однородными условиями существования), локальная популяция (небольшая группа особей, непродолжительное время существующая относительно изолированно) и др.
|
|
|
Однако в литературе до сих пор иногда встречаются и заведомо ошибочные контексты употребления термина «популяция», что наиболее часто бывает связано с подменой этим термином понятия «часть популяции». В результате встречаются, например, выражения типа «размножающаяся популяция оленей» вместо «та часть популяции, которая участвует в размножении» и т. п.
В рядах иерархически соподчиненных биологических систем, отражающих различные уровни организации живой материи, популяция занимает как бы двойственное положение. С одной стороны, она входит в иерархический генетикотаксономический ряд, отражающий филогенетические связи таксонов разного уровня как результат эволюции разных форм жизни. В этом ряду популяция выступает в первую очередь как форма существования вида, занимая место между организмом и видом. Генетическая специфика популяции – общность и специфичность генофонда – определяет наиболее значимое здесь свойство популяции.
Правда, если в более полном объеме принять во внимание генетический и таксономический аспекты построения такого ряда, нам придется включить в него некоторые дополнительные звенья. Так, внутри популяции потомство одной особи либо пары особей характеризуется более специфичным (и менее разнообразным) генофондом по сравнению с генофондом популяции в целом. В совокупности со своими родителями оно образует «семейную группу», у многих организмов легко вычленяемую на практике благодаря тем или иным признакам (локализация в пространстве, поведение и т. п.). В своей совокупности генофонды «семейных групп» образуют генофонд популяции. Аналогом «семейной группы» у растений могут выступать не только особи, полученные из семян одного растения, но и особи вегетативного происхождения (партикулы, клоны).
С другой стороны, обладающие определенной степенью дискретности совокупности фенотипически сходных популяций, населяющих часть географического ареала вида, представляют собой таксономические единицы наиболее низкого иерархического уровня – подвиды (расы). Именно категория подвида отражает дифференциацию популяций на промежуточных этапах видообразования, а подвидовая структура вида, таким образом, является результатом эволюционной истории популяций, в ходе которой они приобретают специфические сочетания признаков. Это обстоятельство позволяет говорить о подвидах как связующем звене между двумя уровнями организации биологических систем – видовом и популяционном.
Правда, подвиды обнаруживаются далеко не у всех видов животных и растений. С одной стороны, существуют так называемые монотипические виды, не образующие подвидов; с другой – в большинстве случаев наука просто не накопила достаточной информации для объективного выделения подвидов. С учетом сказанного место популяции в генетико-таксономической иерархии может быть отображено следующим рядом:
организм → «семейная группа» → популяция → подвид → вид → род → … .
Одновременно с этим, представляя вид в конкретных условия среды, популяция вступает в трофические и иные отношения с популяциями других видов, т. е. включается в состав конкретного биогеоценоза. Этот подход к популяционным системам отражает функциональноэнергетический ряд различных уровней организации жизни:
организм → популяция → экосистема → биосфера.
В этом ряду популяция выступает в качестве функциональной субсистемы конкретной экосистемы, а ее главная функция определяется местом в трофических цепях.
ХАРАКТЕРИСТИКИ ВЕЛИЧИНЫ И СТРУКТУРЫ ПОПУЛЯЦИИ
Популяция имеет определенную организацию и структуру, которые в той или иной степени поддаются описанию. Организация популяции подразумевает в первую очередь такие ее характеристики, как общая величина, характер распределения особей внутри популяции (включая плотность распределения) и механизмы, обеспечивающие устойчивое поддержание популяции как самостоятельной единицы. Под структурой популяции в широком смысле понимают любые подразделения популяции как единичного целого на связанные в определенном порядке части.
2.1. Величина популяции
Величина популяции может характеризоваться в двух аспектах: по числу особей, составляющих популяцию, и по пространству, ею занимаемому (ареалу популяции). Определить первую величину можно только зная вторую. В сравнительно редких случаях, имея дело с очень небольшими популяциями, их величину можно определить путем простого подсчета особей. Гораздо чаще приходится прибегать к так называемому приему экстраполяции: определив среднюю плотность популяции умножать ее на площадь всего ареала, рассчитывая таким образом приблизительное число особей всей популяции.
Очень часто исследователь, пытающийся определить на практике величину конкретной популяции, сталкивается с ситуацией, когда внутри вида, представленного множеством популяций, бывает трудно узнать границы конкретной популяции. Между тем определение таких границ зачастую представляет и значительный практический интерес в связи с пониманием популяции как единицы управления биологическим разнообразием. В связи с этим в рамках популяционной биологии во второй половине ХХ в. сформировалось особое направление исследований – биохорология, устанавливающая подходы и методы определения границ реальных природных популяций. Многообразие экологических связей и особенности филогенетической истории различных биологических видов определили весьма широкий спектр различий в отношении величины их популяций и степени выраженности их пространственных границ.
В природе не так уж редко встречаются популяции с относительно небольшими по площади и весьма четко очерченными ареалами. К числу таковых относятся, в частности, реликтовые популяции, оставшиеся от распространенной в прошлом флоры или фауны. Например, включенный в Красную книгу Республики Беларусь козелец голый (Scorzonera ruprechtiana) на территории нашей страны сохранился только в одном месте – на крутом склоне холма возле оз. Глубелька (национальный парк «Нарочанский»). Небольшая популяция этого растения, безусловно, давно отделена от других популяций данного вида, локализованных на севере Уральских гор. К числу реликтовых относятся также популяции нескольких видов так называемых ледниковых реликтовых ракообразных, изолированно обитающих в нескольких озерах севера
Беларуси: лимнокалянус (Limnocalanus macrurus), реликтовая мизида (Mysis relicta), бокоплав Палласа (Pallasea quadrispinosa), понтопорея (Pontoporeia affinis). Немало других примеров такого рода нетрудно найти в Красной книге любой страны.
Не представляет особых сложностей и определение границ островных популяций, под которыми подразумеваются не только популяции собственно островов, где в качестве фактора изоляции выступают море или океан. К этому же типу относятся и популяции, ареалы которых четко очерчены сушей. Например, популяции рыб гольцов (Salvelinus) в озерах Сибири могут быть настолько хорошо дифференцированными, что особи из разных озер нередко сильно отличаются друг от друга. Хорошо изолированными ареалами обладают очень многие виды высокогорных животных, характеризующиеся ограниченной подвижностью (жужелицы, моллюски и др.) и относящиеся к альпийской зоне. Некогда единый ареал таких популяций оказался раздроблен на ряд фрагментов, разделенных пониженными межгорными котловинами и реками и вполне подходящих под характеристику островных популяций.
В настоящее время многие большие ареалы популяций раздроблены на ряд островных ареалов из-за деятельности человека. В нашей стране это явление происходит с глухарем (Tetrao urogallus). Из-за неграмотной эксплуатации популяций данного вида охотничьих птиц, а также сокращения площади пригодных биотопов и ряда других причин, ареал глухаря в Беларуси на сегодня представлен серией изолированных друг от друга территорий, разделенных пространствами, на которых этой птицы нет.
На фоне общей низкой численности данного оседлого вида существует реальная опасность генетического вырождения многих его островных популяций. У других организмов популяции, напротив, занимают гигантские по площади ареалы, очертить достаточно точные границы которых непросто. Наиболее показательные примеры здесь демонстрируют животные, характеризующиеся высокой степенью подвижности. Например, у европейской городской ласточки (Delichon urbica) все особи, населяющие Европу и Западную Сибирь, представляют по сути единую популяцию.
Их концентрация на зимовке в экваториальной Африке способствует сохранению единого генофонда, и ласточка, появившаяся на свет и окольцованная в гнезде, например, в предместьях Лондона, на следующий год может гнездиться в Новосибирске.
Однако и у перелетных птиц, совершающих ежегодные миграции на большие расстояния, ареал порой оказывается дифференцирован на ряд самостоятельных популяций. В таких случаях именно разные районы зимовок способствуют поддержанию самостоятельности нескольких популяций, например у красноголовой чернети (Aythya ferina), у которой в пределах Евразии выделяются четыре популяции, зимующие в Западном Средиземноморье, бассейне Нила, Индии и Юго-восточной Азии. При этом, как видно из рис. 1, гнездовые ареалы этих популяций отчасти перекрываются, и в некоторых районах бок о бок могут гнездиться особи двух соседних популяций.
Подобные зоны перекрывания ареалов популяций доставляют объективные трудности при их картировании, создавая впечатление отсутствия дискретности популяции как таковой. Однако изучение объективной картины распределения особей в пространстве с учетом их экологической и фенотипической специфики обычно позволяет наметить достаточно выраженные границы популяций и определить зоны их перекрывания. Определенным подспорьем при решении подобных задач может стать использование таксономической категории подвида. Правда, как уже отмечалось выше, подвиды выделяются далеко не во всех группах организмов. «Традиционным» их выделение является у животных с хорошо исследованной географической изменчивостью – у позвоночных, отдельных отрядов насекомых и некоторых других групп. Подвиды представляют собой группы популяций, объединяемых по набору каких-либо морфологических признаков. Чем более ярко выражены такие признаки (например, вариации рисунка крыла у бабочек, окраски оперения у птиц и т. п.) и чем более явственно привязаны их особенности к конкретным географическим регионам, тем легче выделяются подвиды. Однако и здесь исследователи порой сталкиваются с проблемой перекрывания подвидовых ареалов, в которых особи разных подвидов гибридизируют, создавая фенотипическую картину интерградации («перехода») признаков. Определение границ ареалов подвидов, таким образом, может рассматриваться как промежуточный этап в определении и картировании границ ареалов конкретных популяций.
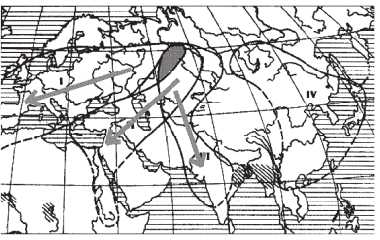
Рис. 1. Популяции красноголовой чернети (Aythya ferina) в Евразии (стрелками показаны направления осенних миграций разных популяций; цветом выделен участок перекрывания ареалов трех популяций в Западной Сибири)
Изучение экологии популяций традиционно велось главным образом на животных; именно в применении к животным были заложены и разработаны существующие подходы в определении границ популяций и подвидов. Однако в последнее время интенсивно стали исследоваться и популяции растений, по отношению к которым ботаники часто употребляют термин «ценопопуляция», который подчеркивает место и роль определенного вида растений в функционировании фитоценоза.
Растительная ценопопуляция включает все особи вида в пределах данного фитоценоза независимо от их фонетического состояния, экотопических и генетических особенностей. Такой подход к характеристике популяций растений определил достаточно четкий принцип выделения конкретных ценопопуляций; их размер и границы определяются границами фитоценоза, которые, хотя и нелинейны, но морфологически выражены более четко, чем границы популяций у животных. Сложности с выделениями границ популяций существуют и в этом случае, но они связаны не со специфическими популяционными проблемами, а с проблемами классификации растительности и методов выделения фитоценозов.
2.2. Пространственное распределение особей в популяции
Внутри ареала популяции особи редко распространяются равномерно. Различают три типа пространственного распределения: случайное (диффузное), равномерное (регулярное) и агрегированное (мозаичное) (рис. 2).
Случайное распределение в природе встречается нередко; в этом случае расстояния между ближайшими особями в популяции различны. Это связано как с неоднородностью среды обитания популяции, так и, в значительной степени, со случайными явлениями. Данный тип распределения часто отмечается у растений и многих животных – в частности у тех, у кого социальные связи между особями выражены относительно слабо. Равномерное распределение характеризуется равным удалением каждой особи от всех соседних особей. В природе такое распределение встречается достаточно редко: на участках с однородной средой обитания, в частности в моновидовых зарослях некоторых растений и плотных поселениях некоторых сидячих беспозвоночных.
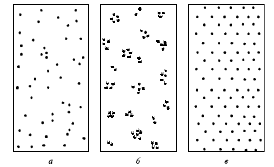
Рис. 2. Типы пространственного распределения особей в популяции: а - случайное; б - групповое (агрегированное); в - равномерное
Агрегированное распределение выражается в образовании группировок особей, между которыми остаются достаточно большие незаселенные участки. Биологически это связано либо с резкой неоднородностью среды, либо с выраженной социальной структурой внутри популяции, как, например, у животных, образующих колонии, либо у растений – потомков вегетативного размножения одной особи.
Пространственное распределение особей часто отражает те или иные особенности состояния популяции: так, при максимально возможной плотности на пригодных участках нередко наблюдается регулярное распределение особей, а если популяция далека от предельной насыщенности – распределение бывает случайным. Агрегированное распределение часто наблюдается, если пригодные для жизни вида участки в пределах ареала популяции «вкраплены» среди непригодных территорий.
Как величина популяции, так и характер распространения особей внутри нее зависят не только от распределения пригодных для жизни условий, но и от биологических особенностей организмов, в первую очередь их подвижности. В биологических исследованиях разных групп организмов нередко возникает необходимость в определении характеристик подвижности особей того или иного вида либо популяции. В частности, исследования перемещений птиц и некоторых других животных с применением мечения и кольцевания позволяют определить среднюю величину дисперсии – среднего расстояния от места появления на свет до места первого размножения особей.
В некоторых случаях удалось выявить даже межпопуляционные различия средней дисперсии в разных популяциях одного вида. Так, исследования обыкновенных поползней (Sitta europaea), населяющих широколиственные леса Центральной Европы, показали, что среднее расстояние между местом появления на свет и местом первого гнездования окольцованных птенцами особей составляет всего лишь около 800 м, а максимальное не превышает 27 км, тогда как у популяций данного вида, гнездящихся в Западной Сибири, эти величины в 2 раза выше.
Однако даже в приведенном примере видно довольно большое различие между величинами средней и максимальной дисперсии, что создает определенные неудобства. Во многих отношениях более показательной характеристикой подвижности особей в популяции является радиус репродуктивной активности (РРА). В настоящее время в популяционной экологии закрепилось понимание этой величины, основанное на эмпирических данных: радиусом репродуктивной активности принято считать расстояние между местом рождения и местом размножения для 95 % особей данного поколения. При исследовании перемещений многих животных было выяснено, что обычно около 95 % особей расселяются на расстояние меньшее, чем половина максимального (т. е. самого дальнего расселения). Разумеется, величины РРА у разных видов сильно варьируют. Ограничимся лишь небольшим рядом примеров:
дрозофилы (Drosophila sp.) разных видов – 150-400 м;
среднеазиатская черепаха (Testudo horsfieldi) –700 м;
зяблик (Fringilla coelebs) – 2 км;
черный коршун (Milvus migrans) – 1000 км;
крот (Talpa europaeus) – 44 м;
лисица (Vulpes vulpes) – 100 км;
песец (Alopex lagopus) – 850 км;
человек (сельские районы Европейской России, конец ХIХ в.) – 10 км.
У некоторых видов РРА подвержен колебаниям у особей разных поколений в разные периоды существования одной и той же популяции, в связи с чем имеют место временные флуктуации ареалов ряда видов, а также нерегулярные дальние миграции (естественные инвазии).
Лишь 5 % особей популяции выселяются за пределы ее РРА. Так называемые «особи-бродяги» иногда уходят очень далеко, порой на расстояния, на порядок превышающие величину РРА. Есть предположение, что склонность к «бродяжничеству» предопределена генетически; в таком случае это явление может иметь эволюционный смысл, обеспечивая потенциал расселения популяции, благодаря которому популяция может расширять свой ареал и осваивать новые территории.
2.3. Плотность популяции
Плотность – величина популяции, отнесенная к некоторой единице пространства; обычно выражается числом особей (реже биомассой) на единицу площади либо объема, например 500 деревьев/1 га, 4 зайца/1 км2, 200 экз. рыбы/1 га водоема и т. п.
Иногда различают среднюю плотность – численность на единицу всего учитываемого пространства – и экологическую, рассчитанную на единицу заселенного данным видом обитаемого пространства (биотопа). Например, если какой-то моллюск в водоеме заселяет только участки с илистым дном, то среднюю плотность придется рассчитывать для всей площади водоема, а экологическую – для суммарной площади заиленных участков.
Иногда на практике более значимым оказывается знание не конкретной плотности популяции в тот или иной момент времени, а наличие возможности оценить ее в сравнительном аспекте – близка она к максимальным показателям или минимальным, возрастает или уменьшается. В подобных случаях полезны показатели относительного обилия. Например, для оценки относительного обилия мышевидных грызунов в исследуемом биотопе выставляют в определенном порядке не менее 25 ловушек и выдерживают их в состоянии готовности не менее 3 полных суток, регулярно собирая «улов». Для сравнения полученных показателей общее число отловленных зверьков пересчитывают к стандартному показателю «число пойманных особей/100 ловушко-суток». Такие показатели по мере их накопления удобны для оценки динамики численности зверьков в разных биотопах, разные сезоны года и разные годы. Оценка абсолютной плотности по этой методике невозможна, и если в ней возникает необходимость, приходится прибегать к гораздо более сложным и трудоемким методам.
Приведенный пример является одним из множества методов оценки абсолютного и относительного многообразия организмов, определяющегося целым спектром особенностей как изучаемых видов организмов, так и биотопов, которые они заселяют.
Подробно рассматривать все существующие методы здесь не имеет смысла; их лучше постигать по мере возникновения надобности, пользуясь специальными методическими руководствами. В приведенном примере с грызунами для проведения исследования приходится прибегать к безвозвратному изъятию зверьков из биотопа; к счастью, это не является обязательным правилом, и большинство методов оценки плотности популяции основаны на подсчете организмов без их умерщвления.
2.4. Возрастная структура популяции
В популяциях большинства организмов можно обнаружить особей разного возраста. Возрастная структура популяции определяется соотношением в ней разных возрастных групп. Эта характеристика существенно влияет на темпы размножения и уровень смертности, а следовательно, на общую численность популяции и ее изменение во времени.
Под возрастной группой понимают группу особей одинакового астрономического либо физиологического возраста. Астрономический (календарный) возраст обычно исчисляют в годах (для недолго живущих организмов – в днях), для выражения физиологического возраста обычно определяют 3 возрастные группы: предрепродуктивную, репродуктивную и пострепродуктивную. При обсуждении различных аспектов возрастной структуры популяций часто используется также термин «поколение», под которым принято понимать непосредственное потомство особей, появившихся на свет на протяжении одного цикла размножения (у однократно размножающихся видов) или всего репродуктивного периода (у видов с неоднократным размножением на протяжении жизни). Впрочем, в отношении человека поколением нередко называют также группу одного астрономического возраста («поколение 2012 года рождения»).
Для выражения соотношения в популяции особей разного астрономического возраста используют так называемые возрастные (демографические) пирамиды (рис. 4; 5). По сути, это диаграммы, на которых число особей каждого возраста (или доля их в популяции) изображено полосой определенного масштаба. Полосы располагаются одна за другой в порядке увеличения значений возраста. Если есть необходимость представить соотношение возрастных групп раздельно по полу, полосы располагают горизонтально, одну над другой по мере увеличения возраста; при этом в левой части пирамиды принято располагать диаграмму для мужчин, в правой – для женщин (рис. 5).
Порой даже беглого взгляда на возрастную пирамиду бывает достаточно, чтобы составить представление об особенностях соотношения воспроизводства и смертности в данной популяции. В нормальной, устойчиво существующей популяции число молодых особей всегда больше, чем число особей старшего возраста; поэтому самой широкой частью такой пирамиды будет ее основание. Однако пирамида с широким основанием, но очень резко суживающаяся кверху (рис. 4, а), чаще всего характеризует популяции с очень высокой смертностью молодняка. Такая ситуация наблюдается, например, у всем известной большой синицы (Parus major): пара этих птиц за весну и начало лета обычно производит на свет два выводка общим числом около 20 потомков, однако до следующего сезона размножения в каждой семье доживают в среднем две птицы.
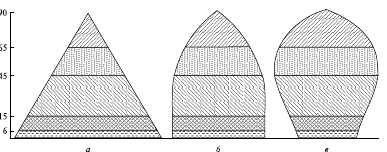
Рис. 4. Демографические пирамиды популяций, построенные без разделения особей по полу: а - растущая; б - стабильная; в - уменьшающаяся
Впрочем, популяция, оказавшаяся в особо благоприятных условиях и быстро растущая, зачастую тоже характеризуется высокой долей молодых особей. Однако по мере замедления темпов роста в такой популяции (с нарастанием ее плотности) либо снизится интенсивность размножения, либо возрастет смертность молодняка, и пирамида изменит свои контуры, приблизившись к варианту, изображенному на рис. 4, б. Однако существует очень мало популяций, у которых такой тип соотношения особей разного возраста в популяции сохранялся достаточно долго.
Есть данные, что близкое к подобному типу соотношение порой наблюдается у некоторых гидр (Hydrida), чья смертность в разном возрасте примерно одинакова. В популяции, численность которой снижается, будет содержаться большая доля старых особей (см. рис. 4, в). Похожий тип пирамид характеризует и некоторые сравнительно устойчивые популяции, если им свойственны высокая выживаемость молодых особей и смертность преимущественно в пострепродуктивном возрасте.
Возрастная структура популяции играет важную роль в демографических и социальных процессах в человеческом обществе. Она тесно взаимодействует с характеристиками воспроизводства человеческого населения. В принципе, все типы возрастных пирамид, отображенные на рис. 4, встречаются и у человека, характеризуя особенности воспроизводства, смертности и тенденции динамики этих характеристик в разных обществах и на разных этапах их развития.
Тип пирамиды, изображенный на рис. 4, а, характеризует страны с высокой рождаемостью и высокой смертностью. У возрастной пирамиды такого населения широкое основание (которое образует высокая доля детей в населении) и узкий шпилеобразный верх (малая доля доживающих до преклонного возраста), боковые стороны имеют вид вогнутых парабол, и чем сильнее вогнуты эти линии, тем выше уровень смертности в детском возрасте. Такой тип воспроизводства населения иногда называют примитивным (по многим параметрам, и не только демографическим, такое наименование ему вполне подходит). В далеком прошлом все народы имели этот тип воспроизводства населения, а некоторые страны сохраняют его и в настоящее время.
Социальное развитие общества приводит к снижению уровня смертности, а спустя 1-3 поколения – и рождаемости. На сравнительно небольшом отрезке времени возрастная структура общества приближается к типу, отраженному на рис. 4, б, а затем, с явным замедлением роста численности населения (о причинах которого пойдет речь ниже), она принимает форму колокола (см. рис. 4, в). Такой тип в демографии иногда называют неподвижным или стационарным.
Процесс снижения смертности рано или поздно замедляется или прекращается (смертность не может сокращаться бесконечно), в то время как снижение рождаемости будет продолжается. Начинается депопуляция — вымирание населения. Самая широкая часть возрастной пирамиды такой популяции постепенно все более смещается вверх, пирамида напоминает форму погребальной урны. Население стареет, т. е. в его составе увеличивается доля пожилых людей и сокращается доля молодежи. Такой тип воспроизводства населения в демографии называют регрессивным или деградационным.
В некоторых случаях возрастная пирамида популяции может иметь форму «елочки», когда длинные и короткие полосы чередуются (см. рис. 5).
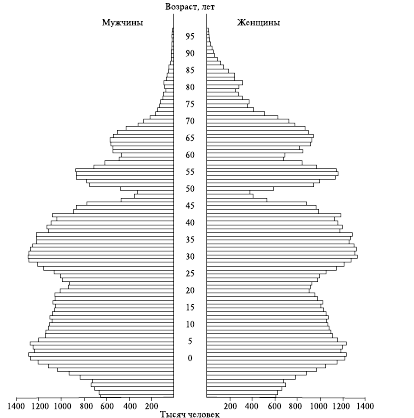
Рис. 5. Демографическая пирамида популяции, построенная с разделением особей по полу
Такой тип характерен для популяций тюленей в местах, где ведется интенсивный промысел недавно родившихся детенышей (так называемого белька), который проводится, как правило, один раз в 2-3 года, но при отсутствии должного контроля, во время которого часто происходит изъятие всего годового приплода.
Для определения физиологического возраста особей выделяют три категории (стадии):
1) предрепродуктивный возраст, когда молодые особи физиологически еще не способны к размножению (у серой крысы, например, до трех месяцев, у воробья до одного года, у гориллы до 5 лет, а у человека до 14 –16 лет);
2) репродуктивный (детородный) возраст, в котором особь способна к размножению (у человека до 45 –50 лет, в виде исключения дольше);
3) пострепродуктивный возраст, когда особь из-за старения организма уже неспособна к размножению.
У многих видов организмов стадия пострепродуктивного возраста отсутствует, например, вплоть до самой гибели сохраняют способность к размножению большинство деревьев, многие птицы. У многих насекомых, а также проходных лососевых рыб, размножение происходит один раз в жизни перед гибелью. В подобных случаях мы можем говорить об очень растянутом предрепродуктивном возрасте при отсутствии пострепродуктивного. У большинства же микроорганизмов выделение групп физиологического возраста на практике невозможно (в микробиологии иногда применяют понятие «возраст колонии микроорганизмов»).
У человека соотношение продолжительности стадий физиологического возраста в самой приближенной форме можно представить как 20:30:20 годам, т. е. в пропорции 1:1,5:1, тогда как у млекопитающих (по крайней мере в условиях неволи) эта пропорция обычно равна 1:2:3. Такое сопоставление породило предположение о потенциальном среднем возрасте человека 120 лет, который не реализуется лишь из-за воздействия неблагоприятных внешних и внутренних факторов.
В популяционной экологии растений принят несколько иной подход к определению возрастной структуры. В составе ценопопуляций выделяют четыре возрастных периода:
1) латентный (семена);
2) прегенеративный (всходы и т. д.);
3) генеративный (размножающиеся растения);
4) постгенеративный (неразмножающие, вплоть до отмирающих).
Ценопопуляции, включающие растения всех возрастных периодов, называются нормальными; они способны к полному самоподдержанию и не зависят от внешнего поступления семян. Популяции же, состоящие преимущественно из особей прегенеративного периода, сильно зависят от заноса зачатков извне. Они характерны для начальных этапов освоения пространства в биоценозе и носят название инвазионных.
Кажущиеся принципиальными различия между особенностями возрастной структуры популяций у растений и животных на самом деле не столь резки, во время исследования колорадского жука Leptinotarsa decemlineata было обнаружено, что часть особей в некоторых популяциях может впадать в состояние затяжной, многолетней диапаузы, а за этот период в активно развивающейся популяции сменится не один десяток поколений. Затем «спящие» жуки по каким-то причинам активизируются и быстро восстанавливают популяционную численность (в том случае, если она, например, была сокращена химическими методами защиты).
2.5. Половая структура популяций
По отношению к животным, подавляющее большинство которых раздельнополы, а также к двудомным растениям (ива, облепиха, тополь и др.), применимо понятие половой структуры популяции, которая определяется численным соотношением самцов и самок в разных возрастных группах. Различают первичное, вторичное и третичное соотношение полов.
Первичное соотношение полов определяется соотношением половых хромосом в процессе мейоза. Обычно наблюдается соотношение 1:1, но нередки и отклонения, которые могут быть обусловлены спецификой определения пола у тех или иных групп организмов, а также полиплоидией и влиянием на половые хромосомы аутосом.
У человека и многих других млекопитающих пол определяется наличием или отсутствием Y-хромосомы. Особи с хромосомными аномалиями (XXY, X0 и др.) в популяциях встречаются редко и соотношение 1:1 нарушают незначительно. Но, например, в популяциях леммингов кроме нормальных ХХ-самок существует некоторое количество ХY-особей, также функционирующих как самки, поскольку локализованный в аутосомах фактор подавляет действие Y-хромосомы. Схожая картина наблюдается у некоторых рыб, например меченосцев.
У многих беспозвоночных (в частности, у Drosophila melanogaster) наличие Y-хромосомы само по себе не определяет пол. Пол этих организмов в основном зависит от численного соотношения определенных хромосом, и отклонения в первичном соотношении полов здесь могут быть гораздо более значительными.
Явление полиплоидии также играет роль фактора, заметно влияющего на первичное соотношение полов. Например, в ряде популяций серебряного карася (Carassius auratus) преобладают триплоидные самки (3n = 160 – 166), а в некоторых регионах (в том числе в Беларуси) самцы у этого вида вообще отсутствуют, и популяции размножаются партеногенетически.
Вторичное соотношение полов — соотношение полов на момент рождения либо ко времени завершения метаморфоза. Отличия вторичного соотношения полов по сравнению с первичным зависят, как правило, от действия факторов внешней среды. У некоторых видов определяющими являются абиотические факторы. Например, у многих амфибий температура воды в период развития икры влияет на соотношение полов у головастиков. А у морского кольчатого червя Bonellia viridis определяющими являются гомотипический биотический фактор: его личинка, если в период свободной жизни ей удастся прикрепиться к взрослой самке, развивается в самца, если же этого не происходит, она оседает на дно и формируется как самка. Другой интересный пример демонстрирует кольчатый червь Ophiriotrocha puerilis, который меняет пол в течение жизни. Молодые особи сначала являются самцами, а когда количество сегментов достигает 15-20, они превращаются в самок.
Иногда вторичное соотношение полов регулируется гормонально. Так, при развитии раздельнополых близнецов у коровы рождаются самец и бесплодная телка-интерсекс, так как мужские гормоны развивающегося самца подавляют развитие половых органов у самки.
Третичное соотношение полов определяется числом самцов и самок среди половозрелых, размножающихся особей, а также особей пострепродуктивного возраста. Отклонения от соотношения 1:1 здесь нередко очень значительны и обычно определяются неодинаковой смертностью самцов и самок в процессе взросления. Поскольку показатели смертности могут существенно не совпадать даже в разных популяциях одного вида, соответствующие различия фиксируются и в третичном соотношении полов.
Для человека и многих других млекопитающих, а также птиц, характерна общая закономерность: уменьшение доли самцов в старших возрастных группах, что обычно связано с более высокой смертностью мужских особей. Эта закономерность хорошо иллюстрируется на примере человеческого населения нашей страны.
Иногда наблюдаются весьма необычные ситуации с третичным соотношением полов. Например, в некоторых популяциях горностая (Mustela erminea) самцы спариваются с еще слепыми, недавно родившимися самками, к этому времени имеющими зрелые яйцеклетки. После такого оплодотворения, хотя и с некоторой задержкой, происходит развитие эмбрионов и рождение вполне нормальных детенышей.
2.6. Этологическая структура популяций
По отношению к животным, характеризующимся более-менее выраженными особенностями поведения, применимо подразделение популяции по признакам поведения входящих в нее особей, т. е. этологической структуры.
Этологические группировки могут выделяться по различным признакам. Наиболее простым является разделение особей на группы по особенностям двигательной активности. Такие группы легко выделяются, например, у ряда видов насекомых, в популяциях которых имеются как крылатые, способные расселяться на большие расстояния формы, так и бескрылые (либо с редуцированными крыльями) особи.
Для ряда высших животных характерно образование более или менее устойчивых «семейных» ячеек из самцов, самок и молодняка (семейные стаи у волков, прайды у львов, стада и табуны у копытных и т. д.). Подобные образования принято объединять под обобщенным названием демы. Поведение особей внутри демов различается: им присущи определенные отношения иерархии, разделение функций между особями и т. п., что вызывает и некоторые особенности в поведении, легко заметные даже при простейших наблюдениях за такими группами. При этом у многих видов наряду с «семейными группами» встречаются и особи, ведущие преимущественно одиночный образ жизни (старые самцы у копытных, неполовозрелые особи у хищных птиц и др.) или же однополые группы не размножающихся особей (группы молодых самцов у морских котиков, стаи неполовозрелых птиц у журавлей и др.). Специфику такой структурированности популяции отражает ее разделение по репродуктивной агрегированности и образу жизни.
С данным подразделением тесно связано деление популяции по поведению внутри групп. Этот подход акцентирует внимание не на характере семейных связей внутри групп, а на особенностях поведения конкретных особей. Исследуя популяцию в таких аспектах, удается выявлять особенности иерархии особей внутри группы, проявления агрессивного поведения, которое может обеспечивать охрану индивидуального участка, влияет на подбор полевых партнеров и ситуации, связанные с конкурентными отношениями за пищу. Нередко при таком подходе удается обнаружить и определенные формы поведенческой специализации отдельных особей внутри группы, например по характеру и способам поиска корма, по тактике избегания хищника и т.п.
Широко известным является разделение популяции по системам брачных отношений. Ключевыми признаками для выявления таких структур является характер брачных отношений и спариваний внутри популяции. Известны следующие системы брачных отношений:
1) моногамия: брачную пару создают один самец и одна самка. При этом пара может образоваться либо на короткий период спаривания (лягушки, многие рыбы, кряква, леопард), либо на один полный цикл размножения (пингвины, аисты, дикие гуси, большинство воробьиных птиц), либо на более длительный период, обычно до гибели одного из партнеров (лебеди, лисы, волки). Гибель партнера в последнем случае не исключает из процесса размножения оставшуюся особь пары, и через какое-то время она находит нового партнера;
2) полигамия: одна особь вступает в брачную связь с двумя и более особями противоположного пола. Различают несколько типов полигамии:
полигиния, когда один самец вступает в брачные связи с несколькими (иногда многими) самками. Видимо, наиболее часто встречающийся тип полигамии у птиц и млекопитающих (африканский страус, крапивник, морские котики, олени и др.);
полиандрия, когда одна самка имеет брачные связи с несколькими самцами. Нередко встречается у рептилий (ящерицы, змеи) и очень редко – у теплокровных животных. Из числа последних наиболее известен пример обитающего в северных тундрах кулика-плавунчика (Phalaropus lobatus), его самка, образовав пару с самцом, спаривается с ним и в построенное за короткий срок гнездо откладывает четыре яйца, после чего покидает своего партнера и ищет нового, с которым спаривание, строительство гнезда и откладка яиц повторяются. Насиживание кладки и вождение выводка целиком ложится на самца. Интересно, что, в отличие от других птиц, самки у этих куликов окрашены более ярко, чем самцы;
полибрахигиния, когда каждый самец спаривается с несколькими самками, а каждая самка – с несколькими самцами. Типичная полибрахигиния наблюдается у птиц-полигамов с хорошо выраженным ритуалом токования (глухарь и тетерев, турухтаны), кукушки, бродячих домашних собак;
промискуитет – структура брачных отношений, при которых любая особь имеет равную вероятность скрещивания с любой другой особью противоположного пола. Типичным примером являются многощетинковые черви и иглокожие, выпускающие свои гаметы просто в море. Определенно сходный тип (но безотносительно к полу) наблюдается и у ветроопыляемых растений. Обычно эти системы брачных отношений причисляют к видовым характеристикам, но в природе они часто сочетаются в одних и тех же популяциях. Например, у многих птиц (выпь, крапивник, дроздовидная камышевка) наряду с самцами-полигамами, на участках которых гнездится по несколько самок, наблюдаются и самцы моногамы, и даже территориальные самцы, остающиеся холостыми. В таких случаях говорят о факультативной полигамии.
Нередко в популяции при детальном наблюдении за входящими в нее особями встречаются определенные предпочтения, связанные с выбором полового партнера.
В таких случаях можно говорить о структуре скрещиваний внутри популяции. Сравнительно редко структура скрещиваний носит случайный характер (панмиксия). Примером панмиктического скрещивания являются иглокожие, выбрасывающие свои половые продукты прямо в воду. Чаще всего наблюдаются так называемые неслучайные структуры скрещиваний.
По генетическому характеру выделяют инбридинг (гомогамию) – скрещивание большей частью генетически схожих особей, и аутбридинг (гетерогамию) — скрещивание генетически различных организмов. Если виду присущ выраженный полиморфизм по каким-либо хорошо заметным внешним признакам, проявления гомогамии или гетерогамии порой можно наблюдать непосредственно в природе. Например, в популяциях азиатского жука-скакуна Cicindela nitida встречаются особи трех типов окраски: зеленые, синие и красные. При этом, наблюдая за жуками в период спаривания, можно видеть, что особи красного типа окраски никогда не спариваются с себе подобными, а только с зелеными либо синими партнерами (пример гетерогамии).
В этологическом плане на структуру скрещиваний часто влияют конкурентные отношения за половых партнеров. В таких случаях преимущество в спаривании получают более конкурентноспособные особи (более сильные, подвижные, ярко окрашенные и т. п.). Однако и это правило не носит абсолютного характера. Например, при чрезмерно высокой численности тетерева (Lyrurus tetrix), когда на ток слетается очень много самцов, самые сильные (и наиболее конкурентноспособные) самцы отвоевывают на токовище центральное место. Прилетевшие на ток самки садятся у его периметра и там спариваются с более слабыми и мелкими самцами, так и не доходя до середины токовища. Возможно, здесь имеет место генетический механизм регулирования численности популяции, вступающий в действие при достижении ей чрезмерно высокой плотности.
Заслуживают внимания и проявляющиеся в популяциях механизмы разнокачественного поведения особей в условиях поддерживаемой иерархии. Так, у некоторых видов грызунов, помимо животных, активно борющихся за свой ранг и поддерживающих его, в группах часто выявляются особи, направленно избегающие схваток, а соответственно, не проявляющие стрессовых реакций. Есть основание полагать, что такие особи имеют большие шансы на успех в размножении. Специальные опыты с мышами показали, что в определенных условиях, помимо доминантов, в размножении регулярно участвуют и другие самцы. Это оказывается возможным потому, что некоторые подчиненные самцы устойчивы к стрессовой ситуации, а у доминантов в критических условиях (борьба за ранг, вторжение «чужака» и т. п.) нарушается сперматогенез, и его восстановление идет медленнее, чем развитие астрального цикла самок.
Интересную ситуацию, связанную со своеобразной структурой скрещиваний и ее последствиями, продемонстрировали опыты М. Смита (1956), поставленные на дрозофиле. Как известно, у дрозофилы каждая самка спаривается лишь один раз и хранит семя в семяприемнике. Самцы же спариваются многократно. М. Смит скрещивал группы самок с двумя линиями самцов – гомозиготной и гетерозиготной, а затем следил за развитием яиц, откладываемых обеими группами самок. Выяснилось, что самки, которые спаривались с гетерозиготными самцами, откладывали столько же яиц, что и другая группа, а вот процент оплодотворенных яиц у них оказался почти в 5 раз выше. Количество спариваний самок с гетерозиготными самцами тоже было гораздо большим, чем с гомозиготными: в течение часа с такими самцами успевали спариться 90 % девственных самок. Оказалось, что самцы обоих типов усиленно ухаживают за самками, но гомозиготные самцы замедленно реагируют на ответный своеобразный танец самок, и многие из самок во время этого промедления улетают, не дождавшись спаривания.
2.7. Генетическая структура популяций
Генетика популяций представляет собой одно из наиболее распространенных направлений популяционной биологии. Природные популяции являются генетически гетерогенными, причем их гетерогенность проявляется в первую очередь различиями аллелей генов у разных особей. Совокупность генов, которые имеются у особей данной популяции, образует ее генофонд. Если конкретные аллели определяют своеобразие генотипа каждой особи, то частоты (концентрация) аллелей и, соответственно, частоты проявления вариантов признаков определяют своеобразие генофонда популяции.
Именно эти явления в первую очередь и отражают генетическую структуру популяции. Генофонд популяции содержит как общие генетические свойства вида, так и особенности, возникшие в порядке приспособления популяции к конкретным условиям ее существования. Именно этот аспект генетической структуры включает степень индивидуальной изменчивости (генетический полиморфизм) по комплексу признаков. С другой стороны, важной особенностью генетической структуры оказывается сложность генома каждой отдельной особи, характеризующаяся степенью гетерозиготности по множеству свойств. Одним из свойств генофонда популяции является его способность накопления и длительного сохранения мутаций (даже неадаптивных) в гетерозиготном состоянии.
Генофонд природных популяций, как правило, не остается стабильным длительное время, так как в природе постоянно действуют факторы, нарушающие генотипическое равновесие. Пример действия таких факторов – естественный отбор, вследствие которого одни генотипы оставляют больше потомков, чем другие, и это вызывает изменение генетической структуры популяции. Второй пример – миграция; из-за притока либо оттока генов генофонд популяции тоже может меняться. Наконец, нередко происходит так называемый генетический дрейф – случайное резкое изменение соотношения частот генотипов в популяции. Такое явление особенно вероятно при резком сокращении численности популяции («эффект бутылочного горлышка»), но порой наблюдается и при стабильной численности популяции. Генному дрейфу, особенно в малочисленных популяциях, сопутствует зачастую инбридинг (близкородственное скрещивание). Эти процессы способствуют снижению уровня генетического разнообразия популяции. Поскольку популяция – элементарная единица эволюционного процесса, процессы преобразования генофонда под давлением отбора и других факторов представляют большой интерес как непосредственные механизмы эволюции.
Однако эти явления в первую очередь считаются предметом исследования генетики популяций. С экологической же точки зрения наиболее интересна специфика эволюционных процессов, происходящих в популяции под воздействием антропогенных воздействий. Это хорошо иллюстрируется примером, полученным при изучении последствий промысла ценной лососевой рыбы – нерки – на Камчатке. У самцов этого вида наблюдается выраженная дифференцированность особей на крупных, но поздно (на четвертом году жизни) созревающих самцов – гомозигот по ряду признаков, и мелких, но созревающих на третьем году гетерозиготных самцов (на Камчатке их называют «каюрками»). В 1930-е гг. доля «каюрок» в большинстве камчатских водоемов не превышала 0,2 % нерестовых стад, а в настоящее время составляет около 40%. В чем же причина столь драматических изменений?
Исследования показали, что главный фактор, определивший столь нежелательное изменение популяции, – избирательный промысел жаберными сетями идущих на нерест крупных гомозиготных самцов. В норме самки нерки проявляют явную избирательность при выборе партнера для нереста, отдавая предпочтение более крупным самцам. Вследствие промысла гомозигот гетерозиготные мелкие самцы получают большие шансы передачи генов следующим поколениям, и их доля в популяции неуклонно возрастает. Подобные процессы происходят в стадах многих промысловых рыб. Сходные последствия наблюдаются и при искусственном воспроизводстве рыбьего поголовья.
Хотя генетические исследования природных популяций в последние десятилетия существенно расширились, все же темпы их развития ограничиваются некоторыми факторами. В частности, генетическое изучение большинства видов живых организмов существенно затруднено сложностью их разведения в неволе и длительными периодами смены поколений.
По этой причине до сих пор не существует ни одного вида животных или растений, у которых была бы изучена генетика всех природных популяций. В определенной степени альтернативным подходом является применение методов фенетики популяций.
Фенетика популяций – распространение генетических подходов и принципов на виды и формы, не изученные генетически, путем исследования фенотипической изменчивости через выделение и рассмотрение дискретных вариаций признаков – фенов. Фенами называют любые дискретные альтернативные вариации признаков и свойств особей, которые на всем имеющемся материале (обязательно многочисленном) далее неразделимы без потери качества. Фены отражают генетическую конституцию данной особи, а своей частотой – генетическую структуру популяции. В одних случаях в качестве фенов выделяются достаточно длинные ряды дискретных вариаций какого-либо признака, и для составления каталога фенов требуется исследовать изменчивость этого признака у большого количества особей. В других же случаях число таких вариаций может быть сравнительно небольшим, и спектр их изменчивости выявляется достаточно легко (например, число дуговидных жилок листа у подорожника (Plantago major), и т. п.
Конечно, предполагать, что каждый фен какого-либо признака соответствует своему состоянию аллели одного гена, определяющего этот признак, было бы чрезмерным упрощением. Нередко оказывается, что под внешне однородным фенотипом у остромордых лягушек (Rana arvalis) на территории Беларуси скрывается действие разных генов.
Известно, например, что фенотипическое проявление мутации black и мутации ebony у дрозофилы практически одинаково – появляются мухи с черным телом. Только генетический анализ позволяет различить эти мутации, локализованные в разных хромосомах. Поэтому экстраполяция знаний из области генетики на генетически неизученные формы (и признаки), конечно же, чревата потерей в точности, но зато позволяет получить существенный выигрыш в широте исследования и является одним из наиболее продуктивных подходов в изучении природных популяций.
Существует два основных направления популяционных фенетических исследований: изучение фенофондов в пространстве, в том числе и для определения естественных границ природных популяций (так называемая феногеография), и изучение структуры и динамики фенофондов во времени. Пример эффективного применения фенетических методов в первом из названных направлений позволил сделать анализ распределения фенов-маркеров позволил не только доказать самостоятельность полесской популяции остромордой лягушки (Rana arvalis), но и достаточно точно определить ареал этой популяции.
В качестве примера применения фенетических подходов во втором из названный направлений можно привести несколько интересных работ, выполненых на основе анализа распределения фенетических признаков листовой пластинки (в частности, числа зубчиков по ее краю) у массовых видов деревьев в зонах длительного техногенного загрязнения. Было показано, что по мере приближения к источнику такого загрязнения (а именно к комплексу никеледобывающих предприятий Мурманской области) в выборках листьев березы увеличивается доля экземпляров с ассимитрией листовой пластинки по названному признаку.
ДИНАМИКА ПОПУЛЯЦИИ
Популяции подвержены постоянным изменениям, даже если неизменными кажутся экосистемы, в которые они входят. По мере того, как вид подстраивается к сезонам года, абиотическим и биотическим факторам среды, меняется величина, плотность, возрастная структура популяции, число родившихся и умерших особей. Изменения в популяции, т. е. ее динамика, представляют основной предмет интереса популяционной экологии. Наиболее разработанный аспект динамики популяций — динамика величины и плотности популяций. Прежде чем приступить к изложению основных закономерностей динамики популяций, следует рассмотреть относящиеся к ней категории, такие как рождаемость, смертность и миграции. Охарактеризуем каждый из этих показателей более подробно.
3.1. Рождаемость и смертность. Кривые выживания
Рождаемость – показатель, отражающий увеличение популяции за счет рождения новых особей. Различают рождаемость абсолютную (физиологическую) и реализуемую (экологическую).
Абсолютная рождаемость – теоретический максимум скорости образования новых особей в идеальных условиях (при отсутствии лимитирующих внешних факторов, когда размножение ограничивается только пределом физиологических возможностей). Экологическая же рождаемость отражает прирост популяции за счет рождения новых особей при фактически имеющихся условиях среды. Если абсолютная рождаемость – величина постоянная, то экологическая – варьирует в зависимости от состава популяции и условий среды.
В некоторых случаях рождаемость можно определить путем прямого подсчета новорожденных в популяции в год. Такой подсчет ежегодно проводится благодаря организованному сбору статистических данных для определения рождаемости человеческого населения в нашей стране. Схожие подсчеты ведутся, например, для поголовья зубров в Беловежской пуще.
Но, даже имея в распоряжении абсолютные цифры рождаемости, для сравнения этих показателей во временном аспекте зачастую имеет смысл приводить их к единому знаменателю. Наиболее часто рождаемость выражают числом родившихся на тысячу особей популяции (без учета пола и возраста) в год. Например, если в популяции, величина которой составляет 100 тыс. особей, за год появилось на свет 5 тыс. новорожденных, то рождаемость в этой популяции составит 50 особей на тысячу («50 промиль»).
У разных организмов рождаемость может исчисляться совершенно различными цифрами - от единиц (например, у китов) до миллионов на тысячу особей (у микроорганизмов, многих рыб, некоторых беспозвоночных). У современного человека рождаемость ниже 15 новорожденных на тысячу человек в год считается низкой, выше 20 на тысячу – высокой.
Нередко рассчитывают и удельную рождаемость как число вновь появившихся особей в единицу времени на одну особь в популяции. И здесь, как в предыдущем случае, удельная рождаемость рассчитывается не относительно числа размножающихся особей, а по отношению ко всем особям популяции. Поэтому при большой доле в популяции особей пострепродуктивного возраста удельная экологическая рождаемость будет ниже.
Смертность – показатель, отражающий уменьшение популяции за счет умирания (гибели) особей. У разных организмов смертность варьируется примерно в тех же пределах, что и рождаемость. Как и рождаемость, она может выражаться общим числом особей популяции, погибших за данный период времени (абсолютная смертность), и, скажем, на тысячу особей в год (удельная смертность). Смертность часто обозначают буквой М (от лат. mors – смерть).
Различают экологическую, или реальную, смертность, – гибель особей в данных условиях среды в данное время, – и теоретическую минимальную смертность, т. е. гибель особей в идеальных условиях, при которых популяция не подвергается лимитирующим воздействиям (т. е. по сути, когда особи умирают только от старости). Возраст, в котором особи умирают от старости, называется физиологической продолжительностью жизни; она зачастую намного превышает экологическую продолжительность жизни.
Доля в популяциях особей, избежавших гибели за какой-то промежуток времени, характеризует выживаемость (если М – смертность, то выживаемость – 1 – М). Смертность (и выживаемость) особей в популяциях наглядно отражаются так называемыми кривыми выживания. Кривую можно получить, если, исходя из совокупного числа новорожденных особей в популяции (т. е. числа всех особей родившегося поколения), отмечать число (или %) выживших в зависимости от времени. Разным популяциям и видам, а также одним и тем же популяциям на разных этапах их существования свойственны кривые выживания различных очертаний (рис. 6). Большинство видов животных и растений подвержены старению, которое проявляется в снижении жизнеспособности с возрастом после периода зрелости.
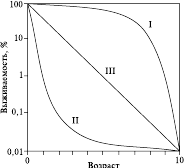
Рис. 6. Кривые выживания
В основе повышения смертности в этом возрасте лежит уменьшение сопротивляемости организма к действию неблагоприятных факторов (например, болезням). Кривая I характеризует те популяции, в которых старение служит главным фактором, влияющим на смертность (например, популяции человека в современной стране с высоким уровнем жизни и развитой медициной). Этот же тип кривой характерен для однолетних растений, особенно культурных, у которых гибель особей на ранних стадиях невелика.
Кривая типа II характерна для популяций организмов с высокой смертностью в ранний период жизни, например для мелких птиц, копытных млекопитающих, а также для человека в странах с низким уровнем жизни и высоким уровнем детской смертности.
Тип III – кривая популяции, имеющей примерно одинаковую смертность в течение всей жизни, когда гибель определяется в основном случайными факторами; характерна для большинства беспозвоночных, многих растений, мышевидных грызунов. Хотя характер изгиба кривых выживания часто очень схож с контуром возрастной пирамиды той же популяции, всегда следует помнить, что кривые выживания показывают смертность одного поколения, тогда как возрастные пирамиды отображают численное соотношение поколений разного возраста в конкретный момент времени.
Вполне понятно, что рождаемость и смертность воздействуют на величину популяции в противоположных направлениях, и увеличение рождаемости обеспечивает увеличение популяции, увеличение смертности уменьшает популяцию. У современных людей смертность в популяции зависит от ее возрастного состава, и чем большую долю в популяции занимают пожилые люди, тем выше показатель смертности. Смертность ниже 15 человек на тысячу считается низкой, выше 20 человек на тысячу – высокой. Один из важных факторов, влияющих на размер популяции, – процент особей, погибающих до репродуктивного возраста. Чтобы численность популяции оставалась стабильной, в среднем только два потомка каждой пары должны доживать до репродуктивного возраста.
Как уже отмечалось, в одной и той же популяции в разные периоды ее существования показатели рождаемости и смертности могут сильно меняться, что в последующем заметно влияет на общую структуру популяции. Это можно легко проиллюстрировать на примере современной структуры человеческого населения Республики Беларусь.
По данным статистики, на 1 января 2013 г. численность населения Беларуси составила 9 млн 463,3 тыс. человек. Если отобразить население на возрастной пирамиде, окажется, что контуры этой пирамиды будут весьма своеобразными и неровными. Причина в том, что населения некоторых лет рождения у нас заметно больше, других – меньше, причем группы таких поколений чередуются, сменяя друг друга. Многочисленными на сегодня являются, например, поколения 1957-1962 и 1982-1987 гг. рождения, а малочисленными – 1971-1975 и 1990-2000 гг. рождения. Причины этого явления имеют историческое объяснение. 1941-1945 гг. – период Великой Отечественной войны, заметно сократившей население страны. Подавляющее большинство мужчин в возрасте от 18 до 45 лет были призваны в армию, в селах и городах не хватало продуктов питания, практически отсутствовала медицинская помощь. Все это стало причиной, с одной стороны, крайне высокой смертности населения детородного возраста, с другой – обусловило весьма низкую рождаемость населения в 1942-1946 гг. Образовались две возрастных группы с очень низкой численностью населения: 1915-1925 и 1942-1946 гг. рождения.
В промежутке между этими «демографическими ямами» оказались поколения 1927-1935 гг. рождения, которые пережили войну с наименьшими потерями и поэтому оказались самыми многочисленными в первое послевоенное десятилетие.
Демографическая статистика показывает, что в среднем наиболее высокую рождаемость обеспечивают поколения в возрасте 25-27 лет. Именно поэтому многочисленные поколения 1927-1935 гг. рождения обеспечили рекордно высокую для послевоенного периода рождаемость в 24 человека на тысячу населения. Такой была рождаемость в Беларуси в 1957-1962 гг., и именно этим объясняется наличие верхнего расширения на сегодняшней демографической пирамиде Республики Беларусь. Поколения 1957-1962 гг. рождения по-прежнему остаются самыми многочисленными в нашей стране.
Из сказанного выше понятна причина последовавшего затем снижения рождаемости – почти до 13 человек на тысячу населения. Оно обусловлено в первую очередь вступлением в детородный возраст самых малочисленных поколений 1942-1946 гг. рождения. Именно поэтому поколения 1971-1975 гг. рождения сравнительно малочисленны, хотя столь сильной «демографической ямы», которая наблюдалась в 1942-1946 гг., здесь уже не отмечается. За счет рождения детей у более старших поколений, а также благодаря иммигрантам происходило некоторое сглаживание контуров демографической пирамиды.
Следующее изменение тенденции рождаемости в нашей стране пришлось на период вступления в детородный возраст многочисленных поколений 1957-1962 гг. рождения. Рождаемость снова стала расти и в 1985 г. почти достигла отметки 18 человек на тысячу населения. Однако эта цифра не сравнялась с рекордным уровнем начала 1960-х гг., что объясняется комплексом причин. Во-первых, как известно, показатели рождаемости в человеческой популяции связаны с возрастом населения, а чем выше средний возраст населения (т. е. чем выше в популяции процентная доля особей пострепродуктивного возраста), тем ниже удельная рождаемость. В 1960 г. население республики было в среднем моложе, чем в 1985 г., в этом и заключается первая причина более низкой рождаемости 1980-х гг.
Вторая причина связана с ростом культурного уровня населения и некоторым изменением психологии людей. Уровень жизни в 1980-е гг. оказался выше, возросли материальные запросы; рождаемость стала более планируемой. Молодые семьи этого периода оказались более расчетливыми, сознательно ограничивая количество детей в семье из-за соображений материального благополучия, а иногда и из-за желания «пожить для себя». Это нормальная тенденция, в той или иной степени проявившаяся в ХХ в. во всех экономически развитых европейских странах. Таким образом, экономика влияет на рождаемость не напрямую, а скорее косвенно, через сознание людей, причем рост материального благополучия, как это не покажется на первый взгляд странным, ведет к снижению рождаемости.
С 1990 по 2000 г. рождаемость в стране вновь падает, причем до крайне низкой отметки 8,5 человек на тысячу населения – демографическое «эхо» последней войны снова напоминает о себе.
В определенной степени контуры демографической пирамиды нашей страны связаны и с таким демографическим показателем, как смертность. В Беларуси смертность населения в настоящее время колеблется около цифр 11 –12 человек на тысячу в год; в периоды спада рождаемости она превышает ее, в периоды подъема оказывается ниже, чем рождаемость. Показатели смертности в долгосрочной перспективе связаны с показателями рождаемости. Например, с вступлением в старческий возраст многочисленных поколений конца 1950-х – начала 1960-х гг. рождения она неминуемо возрастет, и причины этого вполне понятны. С другой стороны, смертность зависит и от таких социальных показателей, как материальное благополучие населения, уровень медицинского обслуживания, образ жизни населения, физическая культура общества. Это следует учитывать при оценке любых статистических данных, касающихся смертности населения в любой стране.
3.2. Миграции
Популяций, величина которых определяется только рождаемостью и смертностью, в природе немного (их называют замкнутыми популяциями). Гораздо чаще встречаются популяции, между которыми существует поток миграции особей. Важно подчеркнуть, что здесь речь идет не о миграциях особей одной популяции внутри ее ареала, а о вселении организмов в популяцию из других популяций (иммиграции) и выселении организмов данной популяции за ее пределы (эмиграции). Вполне понятно, что если особи покидают популяцию (эмигрируют), это приводит к ее уменьшению; если особи вливаются в популяцию (иммигрируют), она увеличивается.
По ряду причин миграционные потоки исследовались в популяциях животных как организмов с выраженной подвижностью особей. В популяциях животных всегда существуют особи, которые размножаются очень далеко от мест рождения, за пределами радиуса репродуктивной активности популяции. Однако именно у животных увеличение популяции за счет иммигрантов ограничивается повышенной смертностью последних. Особи — постоянные обитатели популяции («резиденты»), обычно проявляют повышенную агрессивность по отношению к «чужакам», что часто приводит к их гибели.
Внутрипопуляционные миграции, очевидно, играют роль фактора, повышающего вероятность миграции особей из одних популяций в другие. Так, у популяций ряда видов перелетных птиц гнездовые ареалы географически довольно сильно разобщены, а районы зимовки могут совпадать. Это приводит к дополнительному обмену особями именно в местах зимовки; впоследствии особи, оказавшись в составе «чужой» стаи, вместе с ней мигрируют в ареал «чужой» популяции. Однако такой обмен не должен быть слишком интенсивным, иначе генофонды популяций утратят самостоятельность и популяции сольются.
Другая часть мигрантов покидает границы своей популяции, случайно оказавшись в сфере влияния сильных водных или воздушных потоков, прикрепившись к особям других видов или же попав в грузы, перевозимые человеком. Таким способом на огромные расстояния перемещаются в целом малоподвижные организмы, в том числе растения.
Иммиграция и эмиграция являются значимыми факторами динамики и человеческого населения многих государств, поэтому учитываются соответствующими службами демографической статистики. В частности, наша страна приняла очень большой поток иммигрантов в 1990-е гг., когда происходил массовый выезд белорусов, русских и представителей других национальностей из стран Средней Азии, Кавказа, Прибалтики. С другой стороны, в этот же период происходила довольно значительная эмиграция белорусов из нашей страны в другие государства, которая в значительной мере продолжается и в настоящее время. Учитывать поток эмигрантов довольно сложно, поскольку значительная часть людей выезжает заграницу временно и затем возвращается.
3.3. Рост численности популяции
Исходя из удельных показателей рождаемости, смертности и миграций, можно предложить следующую упрощенную формулу для расчета скорости роста популяции, которую в демэкологии принято обозначать латинской буквой r:
r = [рождаемость + иммиграция] – [смертность + эммиграция].
Из этой формулы видно, что величина r может быть как положительной (популяция растет), так и отрицательной (популяция убывает). Ее называют врожденной скоростью естественного прироста популяции, а иногда – мальтузианским параметром.
Английский священник Томас Мальтус был первым, кто обратил внимание на то, что численность человеческого населения Земли растет в геометрической прогрессии. Скорость роста популяции является мерой удельной (т. е. рассчитанной на одну особь в единицу времени) скорости изменения размера популяции и выражается числом особей в единицу времени.
Максимальная скорость естественного прироста популяции варьирует у разных видов в пределах нескольких порядков; самые высокие значения r наблюдаются у мелких, недолго живущих организмов (например, прокариот). У крупных, с большой продолжительностью жизни (например, человека), максимальные величины r на несколько порядков ниже.
Если условия для роста популяции благоприятны, то скорость роста постоянна и кривая роста имеет вид, изображенный на рис. 7. Такой рост популяции называют экспоненциальным. Если скорость роста популяции постоянна, она зависит от численности популяции N, и чем больше N, тем быстрее растет популяция, и ее численность устремляется к бесконечности:
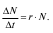
Однако ни одна популяция не может очень долго расти экспоненциально. Со временем каждая популяция сталкивается с теми или иными ограничивающими факторами, которые делают невозможным дальнейший рост (нехватка площади, ресурсов пищи, отрицательное воздействие климатических факторов и др.). Тем не менее, существует целый ряд процессов популяционного роста, который в определенном временном промежутке может быть описан с помощью экспоненциальной модели. Речь идет о случаях нелимитированного роста, когда какая-то популяция заселяет среду с избытком свободного ресурса: одичавшие лошади заселяют пампу, мучные хрущаки – элеватор с зерном, дрожжи – бутыль виноградного сока и т. д.
Однако ресурсы роста популяции исчерпаются и рост затормозится. Каким будет это торможение? В практической экологии существуют разные варианты: резкий взлет численности с последующим вымиранием популяции, исчерпавшей свои ресурсы, постепенное торможение прироста по мере приближения к определенному уровню.
Проще всего описать медленное торможение. Простейшая модель, описывающая такую динамику, называется логистической. Она была предложена для описания роста численности популяции человека французским математиком П. Ферхюльстом еще в 1845 г. В логистической модели вводится переменная K – емкость среды, соответствующая равновесной численности популяции, при которой она потребляет все имеющиеся ресурсы. Прирост в логистической модели описывается уравнением 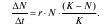
Пока N невелико, на прирост популяции основное влияние оказывает сомножитель r × N, и рост популяции ускоряется. Когда N становится достаточно высоким, на численность популяции начинает оказывать основное влияние сомножитель (K – N)/K, и рост популяции замедляется. Когда N = K, (K - N)/K = 0, рост численности популяции прекращается (рис. 8).
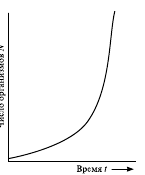
Рис. 7 Кривая экспоненциального роста популяции
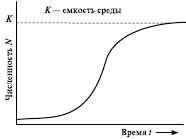
Рис. 8. Кривая логистического роста популяции
Несмотря на простоту, логистическое уравнение удовлетворительно описывает некоторые из наблюдаемых в природе случаев и до сих пор используется в математическом моделировании экологических процессов. Однако гораздо чаще экспоненциальный рост численности переходит значение K, соответствующее реальным экологическим возможностям среды обитания, и еще некоторое время продолжается, более или менее плавно замедляясь. В связи с чрезмерной плотностью популяция обедняет и часто разрушает среду своего обитания, из-за чего вступает в стадию кризиса, в ходе которого численность популяции стремительно падает, в первую очередь путем резкого повышения смертности, чему часто сопутствует снижение рождаемости и увеличение потока эмиграции.
Численность популяции в подобных ситуациях, как правило, опускается ниже того уровня, на котором популяция могла бы существовать стабильно, т. е. входит в стадию демэкологического коллапса. В последующий период среда обитания постепенно восстанавливаются, а вслед за этим растет и популяция. В дальнейшем у разных организмов описанная ситуация либо повторяется снова и снова, либо происходит некоторая стабилизация, и последующие «взлеты» и «падения» оказываются уже не столь значительными (рис. 9).
Факторы, ограничивающие рост популяции, могут зависеть или не зависеть от ее плотности. Практически не зависит от плотности популяции воздействие абиотических факторов (климат, стихийные бедствия и т. п.) и в большинстве случаев – антропогенных факторов. Зависящие от плотности факторы могут действовать с положительной и отрицательной направленностью. Так, смертность в большинстве случаев возрастает с увеличением плотности популяции (положительная, т. е. прямая зависимость), а рождаемость — уменьшается (обратная, или отрицательная зависимость).
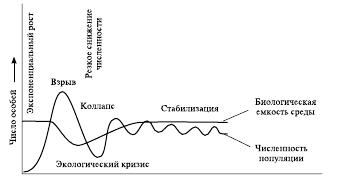
Рис. 9. График колебаний численности природной популяции
Подобным образом действуют на популяцию все биотические факторы: хищники (увеличивается специализация при повышении плотности популяции конкретного вида жертвы), болезни (распространение эпизоотий), конкуренция (как внутривидовая, так и межвидовая), стресс, каннибализм.
3.4. r, K и L стратегии
Периодические нарушения состояния среды («физические стрессы»), такие как пожары, наводнения, ураганы и засухи, часто приводят к катастрофической смертности, внезапно сокращающей плотность популяции до крайне низкого уровня. Популяции многих насекомых быстро растут весной и летом, а с наступлением холодов их численность резко сокращается. Подобные популяции, которые в процессе своего роста дают регулярные или случайные всплески численности, называются оппортунистическими.
Противоположным примером являются равновесные популяции (особенно характерны для многих позвоночных животных) – те, что обычно находятся в состоянии, близком к состоянию равновесия со своими ресурсами, и их плотность гораздо более устойчива (если не меняется количество ресурсов).
Разделение популяций на два типа положено в основу теории стратегий эволюционного развития живых существ. Изначально термин «стратегия» в экологии означал совокупность свойств, помогающих организмам выживать в конкретных условиях, и применялся только по отношению к животным организмам. Р. Макартуром и Э. О. Уилсоном (1967) были выделены r- и K-стратегии по соотношению затрат на размножение и поддержание потомства. K-стратегам (обозначение принято от предельной плотности насыщения популяции K) свойственна низкая плодовитость, в крайнем проявлении – рождение в один сезон размножения всего 1-2 потомков (как, например, у многих крупных млекопитающих и птиц); забота о потомстве, значительная продолжительность жизни. Эти качества направлены на повышение выживаемости во время конкурентной борьбы в условиях длительного существования популяции с высокой плотностью.
r-Стратеги (в связи с обозначением врожденной скоростью естественного прироста популяции r) характеризуются комплексом противоположных свойств: максимальной плодовитостью при отсутствии заботы о потомстве, быстрым развитием особей, что в значительной мере связано с их мелкими размерами; короткой продолжительностью жизни. Типичными r-стратегами являются многие микроорганизмы, насекомые и, конечно же, паразитические черви. Качества r-стратегов направлены на выживание в условиях резко, порой катастрофически меняющейся среды, под прессом не зависящих от плотности абиотических факторов, зачастую – при заселении новых биотопов и новых участков ареала. Однако r-стратеги плохо приспособлены к выживанию в условиях конкуренции.
Р. Уиттекер (1980) различал не два, а три типа стратегий, обозначаемые буквами K, r и L. В основу его системы положены закономерности колебания численности популяций между двумя пределами: K – верхний предел, соответствующий максимальной плотности насыщения популяции, и L – нижний предел, означающий некий «популяционный ноль», соответствующий предельно низкой численности, при которой популяция уже не способна выжить. K-стратеги стремятся к достижению уровня K и длительному существованию в условиях столь высокой плотности, добиваясь этого, в частности за счет предельной дифференциации экологических ниш и выживая в процессе жесткой конкуренции и других взаимодействий в границах занятой среды.
r-Стратеги характеризуются резкими флуктуациями между уровнями K и L. Такие популяции нестабильны и выживают лишь благодаря высокой скорости размножения, они слабо адаптированы как к условиям обостренной конкуренции, так и к неблагоприятным абиотическим условиям среды.
Третья группа популяций – L-стратеги, которые флуктуируют около нижнего предела численности L, хотя иногда могут взрывообразно увеличивать свою численность. У таких популяций отбор имеет тенденцию к совершенствованию механизма для переживания неблагоприятных периодов, а скорость размножения может быть или не быть высокой. Эта схема оказалась применимой не только к животным, но и к растениям.
K-стратеги в мире растений – в основном многолетние виды, часто размножающиеся вегетативно и расходующие в генеративной сфере сравнительно мало энергии. Рудеральные растения, напротив, отличаются укороченным жизненным циклом и высокой семенной продуктивностью, поэтому затраты на размножение здесь больше.
Это соответствует r-стратегии. Группа L занимает переходное положение: например, пустынные многолетники, относящиеся к числу эфемеров с очень быстрым развитием и высокой семенной продуктивностью (результат rотбора), но ежегодно переживающие неблагоприятные условия в вегетативном состоянии (действие K-отбора).
Важно отметить, что выделение r-, K- и L-стратегий – лишь условная модель. В реально существующих природных популяциях выделение «чистых» r-, K- и L-стратегов в большинстве случаев затруднительно. Тем не менее такой подход может быть результативным при сравнении экологических особенностей близких видов, позволяя выявлять существенные различия между их популяциями и объективно оценивать параметры репродуктивной биологии, тенденции динамики численности и т. п. Например, среди синиц (Parus), населяющих лесную зону Европы, при детальном исследовании их биологии выявляются виды, явно тяготеющие к разным экологическим стратегиям. Большая синица (Parus major), по численности в большинстве лесных биотопов доминирующая над другими видами этого рода (хохлатая синица; белая лазоревка), характеризуется высокой интенсивностью размножения (два выводка в год по 10-12 птенцов) и высокой смертностью (r-стратег).
Хохлатая синица (P. cristatus) обладает высоким уровнем специализации в отношении биотопов, выраженной территориальностью и низкими репродуктивными показателями (один выводок в год из 4-6 птенцов) (K-стратег). Наконец, белая лазоревка (P. cyanus), с очень низкой численностью населяющая сильно заболоченные лиственные леса, даже в самых подходящих для нее биотопах характеризуется неравномерным распределением гнездящихся пар. Вместе с тем, благодаря высокому потенциалу размножения, в особо благоприятные годы численность этого вида может резко увеличиваться, следствием чего являются его непериодические миграции («инвазии») далеко за пределы постоянного гнездового ареала (L-стратег).
3.5. Внутрипопуляционные взаимодействия
Организмы внутри одной популяции постоянно воздействуют друг на друга. По своему характеру эти воздействия бывают положительными и отрицательными.
Выражением положительного влияния организмов одного вида друг на друга является групповой эффект – биологические реакции, связанные с объединением организмов одного вида в группы. Достигаемый при этом положительный эффект может быть различным.
У животных довольно часто особи объединяются в группы для облегчения поиска и добычи пищи. Примерами таких взаимодействий являются охотничья стая волков или шеренга, в которую во время ловли рыбы выстраиваются пеликаны.
Очень многочисленны примеры объединения животных в группы для совместной защиты от врагов. Так, гнездящиеся колониями чайки и крачки при появлении вблизи гнездовий пернатого или наземного хищника все одновременно взлетают в воздух и оттуда с громкими криками атакуют неприятеля, обращая его в бегство. В какой-то мере проявлением группового эффекта можно считать и высокую устойчивость по отношению к сильному ветру у деревьев: растущие в лесу с высокой степенью сомкнутости крон деревья в своей массе ослабляют действие сильного ветра и не ломаются под его напором. Иная картина наблюдается после проведения сплошных рубок, когда лес вырубается, но оставляют отдельные так называемые семенные деревья. Последние очень часто ломаются под действием сильного ветра.
С другой стороны, групповой эффект может быть связан еще с некоторыми жизненными функциями, например терморегуляцией. Зимующие у нас мелкие птицы, например корольки и длиннохвостые синицы, проводят морозную ночь, усевшись на одну ветку и плотно прижавшись друг к другу.
Для ряда видов животных и растений характерно взаимообусловленное размножение. Так, многие виды птиц семейства чайковых (озерная и малая чайки, речная крачка и др.) никогда не гнездятся отдельными парами, и одиночные особи либо пары этих видов с наступлением сезона гнездования мигрируют в поисках колоний своего вида или, в крайнем случае, других видов чайковых птиц, и лишь найдя их, приступают к гнездостроению и размножению. Наличие гнездовой колонии в данном случае служит фактором, стимулирующим быстрое развитие половых желез как у самцов, так и у самок.
Примеры взаимообусловленного размножения есть и у растений. Таковым может служить амазонская лилия (Eucharis amasonica) – достаточно известное декоративное луковичное растение. При ее выращивании в цветочных горшках необходимым условием цветения является наличие в горшке нескольких луковиц. Одиночные особи (за очень редким исключением) не цветут.
Следует заметить, что у некоторых видов организмов, в частности у общественных насекомых, условие группового эффекта определяет большинство аспектов жизнедеятельности (добычу пищи, защиту, размножение и др.) и жестко закреплено длительной эволюцией. Групповой образ жизни накладывает отпечаток и на поведение многих общественных животных, что закреплено генетически. Любители певчих птиц знают, что содержание тех же длиннохвостых синиц намного облегчается, если в вольер поместить одновременно несколько особей; в одиночку эта птица в неволе, как правило, ведет себя очень беспокойно и живет недолго.
Массовый эффект, в отличие от группового, характеризуется противоположным, т. е. отрицательным, действием на конкретную особь. Вызывается он в большинстве случаев перенаселением среды. Типичным проявлением массового эффекта у животных является массовый стресс.
Явление стресса в широком смысле слова, согласно автору теории стресса Г. Селье, – неспецифическое биохимическое изменение в организме, вызванное действием любого внешнего фактора, как тривиального (холод, дождь, встреча с другой особью своего вида и т. п.), так и экстремального (опасность, конфликт, травма и пр.).
Массовый стресс — следствие массированного накопления этих изменений у группы особей под действием чрезмерно высокой частоты воздействий тех или иных факторов. Чаще всего он наблюдается в условиях повышенной плотности популяции или же скученности населения.
Физиологические последствия массового стресса у животных могут выражаться по-разному. У многих насекомых (в частности, у мучного хрущака (Tribolium molitor, жук семейства чернотелок)) в условиях повышенной плотности наблюдается удлинение личиночной стадии, что в итоге ведет к снижению плодовитости. У аквариумных рыбок при слишком большом их количестве в аквариуме замедляется рост. Очень частым следствием массового стресса (а у млекопитающих, видимо, неизменным) является повышение агрессивности внутри группы.
Согласно одной из существующих теорий (А. Кребс), массовый стресс является причиной, вызывающей регулярные циклические колебания численности у некоторых млекопитающих, в частности полевок. Экспериментальная группа полевок, содержавшаяся в условиях лаборатории, в достаточном количестве обеспечивалась кормом, водою и всем необходимым. Интенсивное размножение зверьков в этих условиях приводило к быстрому нарастанию плотности (размеры помещения, в котором содержались животные, оставались постоянными). В условиях перенаселенности у животных наблюдалось резкое увеличение надпочечников, что является симптомом сдвига нейроэндокринного равновесия. Следствием этого стало резкое повышение агрессивности и потому – рост смертности, а также снижение плодовитости и уменьшение рождаемости. Конечный результат этого явления – падение численности и уменьшение плотности популяции.
Агрессивность как следствие временного стрессового состояния, вызванного встречей с другой особью своего вида, принадлежит к этой же группе явлений. Она характерна для некоторых высших животных, ведущих одиночный образ жизни. Например, случайная встреча двух землероек-бурозубок (Sorex) вне периода размножения в большинстве случаев влечет за собою яростную драку, которая часто заканчивается гибелью одной, а иногда и обеих особей. Следует заметить, что данная ситуация не связана с защитой территории – землеройки не принадлежат к числу видов с выраженной устойчивой территориальностью. Физиологически явление связано с мощным выбросом адреналина в кровь животного, который стимулируется зрительным образом особи своего вида.
Понятие массового стресса в ряде случаев вполне применимо и для объяснения агрессивного либо немотивированного поведения человека в условиях скученности, например в переполненном общественном транспорте, военных казармах, местах изоляции и др. Психологи достаточно давно заметили негативное влияние массового скопления людей («толпы») на психику индивидуумов.
Очень распространенный пример проявления массового эффекта в популяции – внутривидовая конкуренция, которая определяется как система отношений между особями одной популяции, вызванных недостатком тех или иных потребляемых ресурсов.
Эти отношения у животных могут проявляться в форме эпизодических конфликтов, территориального поведения, систем социальной иерархии, экологической дифференциации особей внутри популяции; у растений – в форме вынужденного ускорения роста, затенения одних особей другими, усиленного ветвления корней, их срастания, образования экотипов. Каждую из этих форм необходимо обсудить более подробно.
Внутривидовая конкуренция происходит между особями популяции одного вида.
Результатом ее является снижение скорости потребления ресурса в расчете на одну особь, понижение скорости индивидуального роста или развития или уменьшение количества запасных веществ в организме. Все это в свою очередь снижает выживаемость и (или) уменьшает плодовитость, которые совместно определяют успех размножения особи. Конечным результатом конкуренции является уменьшение вклада в следующее поколение, т. е. уменьшение по сравнению с тем вкладом, который мог быть сделан в отсутствие конкурентов.
Характерная черта внутривидовой конкуренции – то, что конкурирующие особи разнокачественны генетически и по разному реагируют на недостаток одинаковых ресурсов, что является основой для возникновения асимметричной конкуренции. Существует много примеров, когда при внутривидовой конкуренции наблюдается хорошо выраженная асимметричность: сильный ранний проросток затеняет низкорослый экземпляр, появившийся позднее, а более взрослая и крупная мшанка в прибрежном биотопе перерастает (точнее, обрастает) более молодую и мелкую колонию. Следует также ожидать, что генетически высокорослый злак, как правило, затеняет и подавляет генетически низкорослое растение того же вида. Внутривидовая асимметричная конкуренция может существенно влиять на различия между индивидуальными массами особей в пределах популяции, смещая кривую распределения масс тела влево.
Это свидетельствует о том, что усиление конкуренции приводит к увеличению числа мелких и уменьшению доли крупных особей популяции.
Неравноценность конкурентов означает, что конечный результат конкуренции далеко не одинаков для разных особей. Слабые конкуренты могут внести лишь небольшой вклад в своих потомков или же вовсе не оставить потомства. Вклад сильных конкурентов в потомство может остаться практически неизменным. Другими словами, хотя конечным результатом конкуренции и является уменьшение числа потомков, это не означает снижения индивидуальной приспособленности, особенно для самых сильных конкурентов. Следовательно, было бы неправильно говорить о том, что конкуренция «вредно влияет» на всех конкурирующих особей. В данном случае можно сделать противоположный вывод: сильный конкурент будет становиться все более сильным, а слабый – более слабым.
Характерной чертой внутривидовой конкуренции является также и то, что ее возможное влияние на любую особь тем сильнее, чем большее число конкурентов взаимодействует. Поэтому результаты внутривидовой конкуренции (рождаемость, смертность, рост) следует считать зависимыми от плотности. Продемонстрируем это на примерах.
Эпизодические конфликты, вызванные конкуренцией, проявляются у животных по отношению друг к другу в различных ситуациях – при поедании пищи, борьбе за полового партнера (например, турнирные бои самцов оленей в период гона), выборе места гнездования у птиц (драки скворцов за новый скворечник). Конфликтную агрессивность как проявление внутривидовой конкуренции следует отличать от «стрессовой» агрессивности (хотя биохимическая природа этих явлений общая). В первом случае всегда присутствует явный повод – объект, за который идет борьба (пища, половой партнер и др.), тогда как в условиях массового стресса агрессия часто выглядит немотивированной. Последствия агрессивности при конкурентной борьбе в зависимости от ситуации бывают различными – установление иерархии, определение границ территории, иногда – гибель менее конкурентоспособных особей.
Следует сказать, что природа агрессивности у человека не сводится к стрессовой и конкурентной формам агрессии. Как биологический и социальный феномен, человек обладает сложной гаммой форм поведения, обусловленной рядом факторов. Истоки агрессии человека могут находиться в унаследованных или приобретенных нарушениях психики или же иметь причиной мотивы социального плана.
Территориальное поведение у животных состоит в том, что особь, пара или группа особей (стая, прайд, гарем) занимает определенную территорию, используемую для добычи корма или размножения. Присуще в основном птицам и млекопитающим; изредка встречается у рыб и рептилий (колюшка; крокодилы). Даже среди млекопитающих четко выраженная территориальность свойственна далеко не всем видам и отрядам (практически отсутствует, например, у многих китообразных).
Значение территориальности заключается в том, что особи, занимающие индивидуальный участок, обладают более высокой приспособленностью, чем особи не имеющие его. Они хорошо ориентируются на своей территории, знают расположение ее пищевых ресурсов, места для укрытия от врагов. Действительно, те представители территориальных видов, которым не удалось занять индивидуальный участок, часто не вносят никакого вклада в следующее поколение. Любой выигрыш, который особь получает от наличия у нее индивидуального участка, следует сопоставлять с затратами на защиту этого участка. У некоторых животных защита территории включает жестокие конфликты между конкурентами, тогда как у других существует более тонкое взаимное распознавание конкурирующими особями запрещающих сигналов (например, песня или запаховые метки). Естественный отбор закрепляет такие механизмы поддержания территориальности, при которых выигрыш всегда должен превышать затраты на нее энергии.
К типично территориальным видам относятся хищные птицы (за немногим исключением), совы, ворон, бобр, лиса, выдра, лев, тигр, бурый медведь и др. Эти виды держатся на занятой территории круглый год, изгоняя с нее вторгающихся других особей своего вида; границы территории при этом часто помечаются пахучими секретами особых желез, мочой или другими специальными метками (например, царапинами когтей на коре деревьев у медведя). У птиц своеобразным «маркером» территории часто служит песня. В ряде случаев территория ограничивается лишь местом размножения, а поиск и добыча корма ведутся совместно на общем пространстве. Тогда размеры охраняемой территории могут быть невелики. Так, на лежбище морских котиков территория одного взрослого самца с гаремом занимает лишь 20-30 м пляжа, а пара гнездящихся в колонии чаек охраняет от соседних особей лишь площадку в радиусе 15-20 см от гнезда. У многих видов территория охраняется лишь в период размножения, а позже семья начинает вести кочевой образ жизни. Так ведет себя большинство перелетных птиц, песец.
Социальная иерархия – система поведенческих связей между особями в группе, регулирующая их взаимоотношения (доступ к пище, половому партнеру, убежищу и т. п.) и находящая выражение через доминирование одних особей над другими: первоочередной доступ к пище, водопою, занятие лучших мест в убежище, преимущественное участие в размножении. Ярко выраженные системы иерархии наблюдаются в группах некоторых видов птиц (тетерева, галки, синицы) и многих млекопитающих (крысы, собаки, гиены, львы, горные козлы, большинство обезьян и др.).
Наиболее простой является так называемая линейная форма иерархии (особь А доминирует над особью Б, Б над В и т. д.). Но отклонения от этой схемы бывают очень часто (так называемые сложные формы иерархии). Например, в присутствии особи А особь Б может доминировать над особью В, а в отсутствие особи А наблюдается обратная картина. Подобное нередко имеет место в группах человекообразных обезьян и стаях гиеновых собак, где вожаки-доминанты, занимающие верхнюю ступень иерархической «лестницы», весьма агрессивны по отношению к особям, стоящим на ступень ниже, и гораздо более терпимы к особям самого низкого иерархического уровня.
В обоеполых группах порой устанавливаются отдельные линии иерархии между самцами и самками (у диких, а также одичавших домашних собак). Разумеется, системы иерархии динамичны; со старением и ослаблением доминирующих особей их место занимают окрепшие молодые, а бывший вожак стаи, состарившись, может оказаться на самой низкой ступени иерархической лестницы.
Нарушение системы иерархии часто наблюдается при изменении состояния популяции (в частности, уменьшении либо увеличении численности). Так, у тетерева в условиях невысокой численности преимущество в размножении в период тока имеют более сильные, доминирующие самцы. Если же численность этих птиц велика, на токовище скапливается много самцов. Наиболее крупные и сильные старые петухи в этом случае занимают центр токовища, а прилетевшие самки присаживаются у края и оплодотворяются здесь преимущественно более мелкими и слабыми, обычно молодыми самцами.
Внутрипопуляционная экологическая дифференциация также является одним из следствий внутривидовой конкуренции. Выражается она в частичном разделении экологических ниш между особями одной популяции. Например, в синичьих стаях некоторые особи одного и того же вида предпочитают искать корм в кронах деревьев, другие – в подлеске. У многих хищников более мелкие особи охотятся преимущественно за мелкой добычей, а крупные – за более крупной. В целом это явление распространено в природе достаточно широко, но изучено относительно слабо.
Внутривидовая конкуренция у растений происходит в первую очередь за свет, воду и минеральное питание. Ускорение роста и затенение одних растений другими являются прямым следствием внутривидовой (как и межвидовой) конкуренции растений за свет. В результате затенения у менее конкурентоспособных особей снижается фотосинтез, следствием чего является редукция корневой системы, снижение интенсивности транспирации, подавление микоризообразования и затруднение в поглощении фосфора. Растение в этом состоянии более подвержено действию вредителей и болезней и в конце концов погибает.
Конкуренция за водное и минеральное питание имеет следствием интенсивное ветвление корневой системы, а у деревьев еще и срастание корней. Последнее явление в природе распространено достаточно широко. В старых насаждениях сосны, например, оказываются сросшимися корнями 25 –30 % деревьев, а у дуба порой это число достигает 100 %. В таких случаях корневая система даже срубленного дерева живет несколько лет, получая питание от корней соседних деревьев.
Формирование экотипов (генетических линий, адаптированных к разным экологическим условиям) у растений в большинстве случаев связано с существованием в различных местообитаниях. Однако в некоторых случаях это явление, очевидно, может быть и следствием внутривидовой конкуренции (например, формирование рано- и поздно-цветущих линий внутри одной популяции).
Здесь мы не останавливаемся подробно на эволюционной роли внутривидовой конкуренции как одной из важнейших форм борьбы за существование. Ее действительные эволюционные последствия крайне многогранны. Лишь в качестве примеров можно назвать происхождение деревьев как жизненной формы; появление растений с ярким крупным околоцветником; формирование устойчивых типов брачных отношений у животных; развитие полового диморфизма и связанного с ним разделения экологических ниш, и многое другое.
3.6. Колебания численности популяции
Колебания численности являются неотъемлемым свойством всех популяций. Особенно заметны они у животных и микроорганизмов; несколько сложнее оценивать эти колебания у растений, так как здесь приходится учитывать и факт существования специализированных стадий — спор и семян, что крайне затрудняет даже приблизительный подсчет числа особей в какое-либо время.
Диапазоны колебаний численности у различных животных варьируются в очень широких границах. Если, например, у рыси минимальная и максимальная численность равна 1:10, то у насекомых-фитофагов это соотношение может составлять 1:10 000 000.
Наиболее хорошо заметны так называемые сезонные изменения численности, связанные с сезонными колебаниями факторов среды (например, быстрое размножение насекомых-фитофагов летом и их массовая гибель от осенних заморозков, и т. п.). Как правило, популяции, подверженные подобным изменениям численности, обладают комплексом адаптаций, обеспечивающих выживание в меняющихся условиях (например, откладывают яйца до наступления холодов).
Вторую группу колебаний численности популяции представляют ее случайные изменения, не носящие периодического характера и не связанные с сезоном года. Причины таких колебаний чаще связаны с биотическими факторами среды, такими как доступная пища, болезни и т. п. У некоторых видов изменения численности носят настолько регулярный характер, что к ним вполне применим термин «циклы». Классическим примером 9-10–летних циклических изменений служит изменение численности зайца-беляка и рыси, открытые Ч. Элтоном путем изучения отчетов Компании Гудзонова залива по заготовкам пушнины. Оказалось, что в Канаде и на Аляске с 1736 г. пики численности у рыси повторяются каждые 9-10 лет: они обычно сменяются резкими спадами, когда на протяжении 2-3 лет рысь встречается крайне редко, а затем ее численность вновь растет.
Такие же циклы наблюдаются у зайца-беляка, но его пики численности, как правило, на год или два предшествуют пикам рыси. Очевидно, что цикл хищника в данном случае связан с циклом жертвы, но интересно, что такая же цикличность колебаний численности зайца отмечена и в тех районах, где рысь вообще не водится.
Для тундровых грызунов леммингов зафиксированы трех- или четырехлетние циклические колебания численности; с ними связаны и циклы основных хищников, поедающих лемминга, – песца, полярной совы. В Скандинавии в определенный период численность леммингов становится настолько высокой, что происходит их массовая миграция из перенаселенных местообитаний, совершаемая обычно в северном направлении. Орды мигрирующих грызунов порой достигают моря и тонут.
Какие же причины вызывают подобные циклы? Очевидно, «универсальной» причины не существует. Существует ряд гипотез (далеко не всегда взаимоисключающих), которые в той или иной степени удовлетворительно объясняют это явление у определенных видов.
1. Гипотеза колебаний в системе «хищник – жертва», суть которой в следующем.
Массовое размножение каких-то организмов, составляющих основу питания хищников, приводит к массовому размножению последних. Численность хищников вырастает настолько, что они уничтожают подавляющее большинство особей жертв, после чего сами либо начинают погибать от голода, либо мигрируют в другие регионы.
В итоге немногие оставшиеся в живых особи жертв, не подвергаясь прессу хищников, беспрепятственно размножаются и восстанавливают свою первоначальную высокую численность. Вслед за этим следует размножение (или иммиграция) хищников, и все повторяется вновь.
Подобным образом, в частности, объяснял выявленные им популяционные циклы у зайца и рыси Ч. Элтон. Следует заметить, что циклические колебания такого рода сильнее всего выражены в менее сложных экосистемах с малым числом видов хищников и их потенциальных жертв, например в экосистемах северных тундр.
2. Гипотеза количества пищи. Нередко рассматривается как частный случай предыдущей гипотезы. С ее помощью обычно объясняют популяционные циклы, наблюдаемые на низких уровнях трофических связей (первичные продуценты/консументы I порядка). Впервые она была предложена для объяснения 10-летних циклов у зайца беляка. При высокой численности зайцев заметно сокращаются ресурсы доступной пищи (зеленой массы кустарников и травянистых растений), что приводит к сокращению числа зайцев. С восстановлением растительности увеличивается и плотность популяции зайцев.
В ряде случаев циклические колебания плотности популяций видов-консументов напрямую связаны с циклами растений-продуцентов, служащих для них источником корма. Так, наблюдаемые один раз в 4-5 лет обильные «урожаи» семян ели всегда сопровождаются возрастанием численности их потребителей – рыжих полевок и желтогорлых мышей.
3. Гипотеза стресса. Основывается на медицинской теории стресса, созданной Г. Селье. В середине ХХ в. были получены данные о природных и лабораторных популяциях полевок, у которых в условиях перенаселенности происходит увеличение надпочечников. Это – один из симптомов сдвига нейроэндокринного равновесия, который приводит к резкому повышению агрессивности и снижению (а в крайних случаях — полному прекращению) размножения, что влечет за собой падение численности. А. Кребс (1970) отразил эту закономерность в виде схемы.
Увеличение плотности популяции
↓
Усиление «стрессорного напряжения»
↓
СТРЕСС
↓
Уменьшение интенсивности размножения
↓
Увеличение смертности
↓
Уменьшение плотности популяции
↓
Снятие «стрессорного напряжения»
↓
Увеличение плотности популяции
Заметим, что в условиях «стрессорного напряжения» агрессия может выступать как прямой фактор ограничения численности. Это, в частности, было хорошо показано в экспериментах с домовыми мышами. В искусственных популяциях данных животных рост численности и плотности населения всегда сопровождался увеличением числа драк между особями. По достижении определенного порога частота драк становилась фактором ограничения дальнейшего роста плотности в первую очередь из-за высокой гибели молодых особей.
Специфическая форма блокирования размножения, описанная у ряда видов мелких грызунов, заключается в прерывании беременности в присутствии «чужого» самца, т.е. в ситуации, естественно учащающейся при повышении плотности населения в природных условиях. В экспериментах с серыми полевками, леммингами, домовыми мышами содержание беременных самок с незнакомыми им самцами в 55-87 % случаев ведет к прерыванию беременности. Механизм этого явления, названного «Брюсс-эффект» (по имени описавшей его исследовательницы), связан именно с развитием стресса: у подопытных самок регистрируется повышенный уровень свободных кортикостероидов в крови.
Не менее важной причиной является увеличение смертности в силу повышения чувствительности стрессированных животных к влиянию неблагоприятных факторов среды. При такой ситуации условия, которые в обычном состоянии воспринимаются как нормальные, при повышении уровня стресса оказываются для животных губительными.
4. Гипотеза генетического контроля. Предположение о генетической основе популяционных циклов впервые высказали В. Г. Веллингтон (1957) и Д. Читти (1960). Первый из авторов изучал популяции коконопряда и заметил, что на подъеме численности гусеницы коконопряда плетут длинную паутину, активно питаются и перемещаются («сильные поколения»). В пике же численности гусеницы менее активны, плетут короткую паутину, меньше едят и более восприимчивы к болезням («слабые» поколения).
Было высказано предположение, что в популяциях коконопряда сформирован контролирующий генетический механизм, благодаря которому предотвращается полная деформация и гибель кормовых деревьев, что вызвало бы гибель и насекомых.
Несколько иную генетическую интерпретацию возможных механизмов цикличности предложил Д. Читти, изучавший динамику популяций полевок. Согласно его гипотезе, в условиях повышенной плотности популяции усиливается действие конкуренции между особями, что «включает» действие отбора по K-стратегии. Это приводит к изменению генофонда популяции, однако наступающая депрессия численности устраняет фактор отбора «на конкуренцию», и в последующих поколениях наблюдается сдвиг соотношения генотипов в обратном направлении (в сторону r-стратегии).
Поскольку генетическая реакция популяции всегда отстает от изменения давления отбора, устойчивое равновесие никогда не устанавливается.
Существуют и другие, еще более частные гипотезы. Следует сказать, что, как правило, не выдерживают испытания факторами гипотезы, пытающиеся связать популяционные циклы с абиотическими факторами среды (солнечными пятнами, климатом и т. п.).
Дата добавления: 2022-06-11; просмотров: 38; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!