Тема: Статистичний характер законів спадковості та їхні цитологічні основи.
Мета: Познайомити студентів із явищем щепленого успадкування. Дати чітке уявлення про генотип як цілісну систему та генетику статі. Розвивати увагу, вміння сконцентруватися. Сприяти формуванню пізнавального інтересу. Виховувати зацікавленість дисципліною.
Методи: словесний, наочний.
План:
1 Хромосомна теорія спадковості.
2 Генетика статі.
3 Цитоплазматична спадковість.
Матеріально-технічне забезпечення та дидактичні засоби, ТЗН: підручник, таблиці, малюнки.
Література:
1 Тагліна О.В. Біологія, 10 клас (рівень стандарту, академічний рівень). Підруч, для загальноосв. навч. закл.- Х.: Вид-во «Ранок», 2010. – 256 с.
2 Кучеренко М. Е., Вервес Ю. Г., Балан П.Г. та ін.. Загальна біологія, 10- 11 класи.- К.: Генеза, 1998, 2000, 2001.
3 Кучеренко М.Є. Загальна біологія. Підручник для 10 кл. Загальноосв. навч. закл. – К.: Генеза, 2005 р.
Що таке закон чистоти гамет? На той час, коли Мендель проводив свої досліди з горохом, ще нічого Не було відомо про гени, будову хромосом і процес мейозу. Але він запропонував закон чистоти гамет, який згодом знайшов своє експериментальне підтвер- дження. Цей закон стверджує, що в гібридного (гетерозиготного) організму гамети «чисті». Тобто кожна з гамет такого диплоїдного організму може мати лише один алельний ген і не може одночасно нести дві алелі.
Які цитологічні основи і статистичний характер законів спадковості? Нестатеві (соматичні) клітини, як правило, мають диплоїдний набір хромосом, тобто в кожній з них є два алельні гени. Це можуть бути дві домінантні чи дві рецесивні алелі певного гена {гомозигота) або домінантна і рецесивна алелі {гетерозигота). Коли внаслідок мейотичного поділу утворюються статеві клітини, в кожну з них потрапляє тільки одна алель з кожної пари. Гомозиготна особина формує лише один сорт статевих клітин (домінантною або рецесивною алеллю певного гена), тоді як гетерозиготна - два сорти в рівних кількостях (50% з домінантною алеллю певного гена і 50% — з рецесивною).
Простежимо за гомологічними хромосомами при моногібридному схрещуванні гомозиготних особин гороху посівного. Для спрощення припустимо, що такі особини мають лише одну пару гомологічних хромосом (тобто кількість хромосом у диплоїдному наборі дорівнює двом: 2п = 2), а кожна з них містить тільки один ген. Хромосому з домінантною алеллю (А) на малюнку позначено жовтим кольором, а з рецесивною (а) - зеленим. Відомо, що нащадки, отримані від схрещування особин гомозиготних за домінантною та рецесивною алелями (гібриди першого покоління), будуть гетерозиготними (їхній генотип — Аа). Це пояснюється тим, що одну хромосому з домінантною алеллю вони дістають від одного з батьків, а іншу, з рецесивною, -від другого. Отже, такі рослини будуть одноманітними як за генотипом, так і за фенотипом.
Гібриди першого покоління, на відміну від батьків, утворюватимуть гамети двох сортів: половина з них нестиме хромосому з домінантною алеллю, половина - з рецесивною. Внаслідок схрещування гібридів першого покоління між собою у їхніх нащадків (гібридів другого покоління) можливі три варіанти генотипів: чверть особин матиме хромосоми лише з домінантними (гомозиготи за домінантною алеллю - АА), половина — одну хромосому з домінантною, другу — з рецесивною (гетерозиготи — Аа) і чверть — хромосоми лише з рецесивними (гомозиготи за рецесивною алеллю — аа) алелями. А за фенотипом три чверті насіння гібридів другого покоління матиме жовте забарвлення (гомозиготи за домінантною алеллю і гетерозиготи), а одна чверть - зелене (гомозиготи за рецесивною
алеллю).
Отже, якщо утворюється значна кількість гамет рівної життєздатності, то стає зрозумілим статистичний характер закону розщеплення. Він визначається рівною ймовірністю зустрічей гамет різних сортів. Результати проведеного Г. Менделем моногібридного схрещування можна записати схематично. 
Проте простіше записати хід схрещування за допомогою так званої решітки Пеннета*. її зображують так: по горизонталі записують гамети однієї (у роздільностатевих організмів — батьківської) особини, а по ветрикалі — іншої (материнської). В місцях перетину горизонтальної та вертикальної ліній зазначають генотипи нащадків {схема 2) 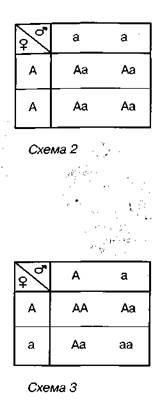 .
.
Згідно з записом, усі нащадки, одержані від схрещування особин, гомозиготних за домінантною і рецесивною алелями, є одноманітними й гетерозиготними.
Результати схрещування між собою гібридів першого покоління відображені на схемі 3.
Отже, ми бачимо, що серед гібридів другого покоління можливе утворення трьох варіантів генотипу. За умови повного домінування домінантної алелі над рецесивною вони визначатимуть два варіанти фенотипу. З даних схеми стають зрозумілими і причини подальшого розщеплення ознак унаслідок розмноження гібридів другого покоління самозапиленням. Гомозиготні особини будуть формувати гамети лише одного сорту, тому серед їхніх нащадків не спостерігається явище розщеплення. Гетерозиготні особини утворюватимуть гамети двох сортів (з домінантною і з рецесивною алеллю), тому серед їхніх нащадків буде спостерігатися розщеплення за фенотипом у співвідношенні 3:1.
Так само можна продемонструвати цитологічні основи і статистичний характер закону незалежного комбінування станів ознак. Уявімо, що в диплоїдному наборі рослина має дві пари гомологічних хромосом (2п = 4), кожна з яких несе лише один гей. Хромосоми однієї пари позначимо паличками, іншої - кружечками. Хромосоми, які несуть домінантну алель, забарвлені у жовтий колір, а рецесивну — в зелений.
Припустімо, що материнський організм містить хромосоми лише з домінантними алелями генів забарвлення і структури поверхні насіння, а батьківський -лише з відповідними рецесивними. Такі гомозиготні організми в процесі утворення гамет формуватимуть один їхній сорт - або з домінантними, або з рецесивними алелями. У разі схрещування батьківських форм усі нащадки (гібриди першого покоління) дістають по дві хромосоми з відповідними домінантними алелями від материнського організму і по дві з рецесивними -від батьківського. Тож усі вони будуть гетерозиготними за генами забарвлення і структури поверхні насіння й формуватимуть чотири сорти гамет у рівних кількостях. При цьому в двох з них алельні гени будуть знаходитися у тих самих комбінаціях, що і в гаметах вихідних батьківських форм, а в двох інших - у нових поєднаннях (рекомбінаціях). Рекомбінація (від лат. ре — префікс, який означає поновлення, повтор дій, і комбінатів - поєднання) - перерозподіл спадкового матеріалу батьків у генотипі нащадків. Іншими словами, рекомбінації - це нові поєднання алелей різних генів у гаметах гібридів, які відрізняються від їхнього поєднання у гаметах батьків.
Унаслідок рівної ймовірності зустрічей гамет різних сортів, які утворюють гібриди першого покоління, в їхніх нащадків (гібридів другого покоління) можливе утворення дев'яти варіантів генотипів. Ці варіанти, у свою чергу, визначатимуть чотири різних варіанти фенотипу. У двох із них стани ознак забарвлення насінини і структури її поверхні будуть у тих самих поєднаннях, як і у фенотипі вихідних батьківських форм (жовтий колір — гладенька поверхня насінини і зелений колір - зморшкувата поверхня), а у двох - в нових - рекомбінаціях (жовтий колір - зморшкувата поверхня і зелений колір - гладенька поверхня). Рекомбінації є одним із джерел мінливості організмів.
Статистичний характер закону незалежного комбінування станів ознак можна довести, записавши хід Дигібридного схрещування за допомогою
решітки Пеннета. При схрещуванні гомозиготних батьківських форм його результат буде такий:
| ab | ab | ab | ab | |
| АВ АВ АВ АВ | AaBb AaBb AaBb AaBb | AaBb AaBb AaBb AaBb | AaBb AaBb AaBb AaBb | AaBb AaBb AaBb AaBb |
Результати схрещування батьківських форм, гомозиготних за домінантними і відповідно рецесивними алелями генів забарвлення (А - а) і структури поверхні (В — Ь) насіння, розташованих у негомологіч-них хромосомах, підтверджують закон одноманітності гібридів першого покоління: всі вони гетерозиготні за цими генами й утворюватимуть насіння жовтого кольору з гладенькою поверхнею. Серед гібридів другого покоління розщеплення за фенотипом буде таким: 9 частин насіння жовтого кольору з гладенькою поверхнею, 3 — жовтого зі зморшкуватою, 3 - зеленого з гладенькою і 1 — зеленого зі зморшкуватою. Отже, розщеплення серед гібридів другого покоління як за ознакою забарвлення насіння, так і за ознакою структури його поверхні становитиме 12:4 (тобто 3:1). За умови повного домінування домінантних алелей над відповідними рецесивними фенотип жовтого насіння з гладенькою поверхнею визначатиметься чотирма варіантами генотипу (ААВВ, ААВЬ, АаВВ, АаВЬ), жовтого зі зморшкуватою - двома (AAbb, Aabb), зеленого з гладенькою - теж двома (ааВВ, ааВЬ), а зеленого зі зморшкуватою - одним (aabb).
Що таке явище зчепленого успадкування? Закон незалежного комбінування станів ознак ґрунтується на таких положеннях:
- розвиток різних станів ознак зумовлений алельними генами, які мають однакове розташування в гомологічних хромосомах;
- гамети та інші гаплоїдні клітини, які мають по одній хромосомі з кожної пари гомологічних хромосом, несуть лише один алельний ген із певної їхньої кількості;
- гени, що контролюють розвиток ознак, які успадковуються незалежно, розташовані в негомологічних хромосомах.
Коли ми доводили статистичний характер законів спадковості, встановлених Г. Менделем, то для спрощення припускали, що кожна хромосома несе лише один ген. Але вчені вже давно звернули увагу на те, що кількість спадкових ознак організмів значно перевищує кількість їхніх хромосом у гаплоїдному наборі. Так, у гаплоїдному наборі мухи-дрозофіли, яка стала класичним об'єктом для генетичних досліджень, усього чотири хромосоми. Але кількість спадкових ознак і, відповідно, генів, які їх визначають, безсумнівно, значно більше.
Отже, в кожній хромосомі є не один, а багато генів. Разом з ознаками, які успадковуються незалежно, існують і такі, що успадковуються зчеплено одна з одною, оскільки визначаються генами, розташованими в одній хромосомі. Такі гени утворюють групу зчеплення. Кількість груп зчеплення в організмів певного виду дорівнює кількості хромосом у гаплоїдному наборі (наприклад, у дрозофіли In = 4, у людини -In = 23).
Експериментально явище зчепленого успадкування довів видатний американський генетик Т.Х. Морган зі своїми співробітниками. Слід відзначити, що цей учений, як свого часу і Г. Мендель, вдало обрав об'єкт для своїх досліджень - муху-дрозофілу. Цих комах легко утримувати в лабораторіях. Вони мають високу плодючість і швидку зміну поколінь (за оптимальних умов утримання нове покоління з'являється кожні півтора-два тижні), невелику кількість хромосом, що значно спрощує проведення досліджень. Явище зчепленого успадкування Т.Х. Морган встановив за допомогою такого досліду. Самців дрозофіли, гомозиготних за домінантними алелями забарвлення тіла (сіре) і форми крил (нормальна), він схрестив із самками, гомозиготними за відповідними рецесивними алелями (чорне забарвлення тіла -недорозвинені крила). Всі гібриди першого покоління мали сіре забарвлення тіла і крила нормальної форми, тобто були гетерозиготними за обома генами. Щоб з'ясувати генотип гібридів першого покоління, Т.Х. Морган провів аналізуюче схрещування: схрестив їх з особинами, гомозиготними за відповідними рецесивними алелями.
Теоретично від такого схрещування можна було очікувати два варіанти розщеплення. Якби гени, що визначають забарвлення тіла і форму крил, були роз ташовані в негомологічних хромосомах, тобто успадковувалися незалежно, розщеплення мало бути таким: 25% особин із сірим тілом і нормальною формою крил, 25% - із сірим тілом і недорозвиненими крилами, 25% - з чорним тілом і нормальними крилами і 25% - з чорним тілом і недорозвиненими крилами (тобто у співвідношенні 1:1:1:1). Але якби ці гени були розташовані в одній хромосомі та успадковувалися зчеплено, то серед нащадків можна було б очікувати 50% особин із сірим тілом і нормальною формою крил і 50% - з чорним тілом і недорозвиненими крилами (тобто у співвідношенні 1:1).
Реально 41,5% нащадків мали сіре тіло і нормальну форму крил, 41,5% - чорне тіло і недорозвинені крила, 8,5% - сіре тіло і недорозвинені крила і 8,5% -чорне тіло і нормальну форму крил. Отже, розщеплення за фенотипами наближувалося до 1:1 (як у випадку зчепленого успадкування). Але серед нащадків були й два інші варіанти фенотипів, які можна було б очікувати в тому випадку, якби гени забарвлення тіла і форми крил містились у негомологічних хромосомах й успадковувалися незалежно.
Як Т.Х. Морган пояснив ці результати? Він припустив, що гени, які визначають забарвлення тіла мух і форму їхніх крил, розташовані в одній хромосомі й успадковуються зчеплено. Під час утворення статевих клітин у процесі мейозу гомологічні хромосоми здатні обмінюватися своїми ділянками (явище перехресту хромосом, або кросинговер). Як вам відомо, кросинговер підсилює мінливість, забезпечуючи утворення нових варіантів поєднань алелей. При цьому може відбуватись обмін кількома генами або ділянками одного гена. Явище кросинговеру згодом було доведено за допомогою введення в різні ділянки гомологічних хромосом ізотопних атомів (міток). Після завершення кросинговерутакі мітки, введені в одну з гомологічних хромосом, виявляли у відповідних ділянках іншої.
Дослідження кросинговеру, проведені на різних організмах, виявили такі закономірності:
- сила зчеплення між двома генами, розташованими в одній хромосомі, обернено пропорційна відстані між ними. Отже, чим ця відстань більша, тим частіше відбувається кросинговер;
- частота кросинговеру між двома генами, розташованими в одній хромосомі, є величиною відносно постійною для кожної конкретної пари генів.
На підставі проведених досліджень зроблено важливий висновок - гени розташовані в хромосомі по її довжині в лінійному порядку, тобто один за одним. Згодом відкриття будови молекули ДНК підтвердило це. Як ви пам'ятаєте, в ній у лінійному порядку розташовані окремі мономери — нуклеотиди, якими закодовано спадкову інформацію.
Хоча частота кросинговеру між різними парами зчеплених генів є величиною відносно постійною, на неї можуть впливати деякі фактори зовнішнього і внутрішнього середовища (зміни в будові окремих хромосом, які ускладнюють або унеможливлюють процес кросинговеру, висока або низька температура, рентгенівські промені, деякі хімічні сполуки тощо). У деяких організмів виявлено залежність частоти кросинговеру від віку (наприклад, у дрозофіли) або статі (миші, кури).
Що таке генетичні карти хромосом? Відносну сталість частоти кросинговеру між різними парами генів однієї групи зчеплення використовують як показник відстані між окремими генами, а також для визначення послідовності їхнього розташування в хромосомі. Зокрема, визначення частот кросинговеру між різними парами генів використовують для складання генетичних карт хромосом. На цих картах зазначено порядок розташування і відносні відстані між генами в певній хромосомі.
Що таке хромосомна теорія спадковості? За результатами своїх досліджень Т.Х. Морган із співробітниками запропонував хромосомну теорію спадковості. За її допомогою з'ясовано матеріальну основу законів спадковості, встановлених Г. Менделем, і те, чому в певних випадках успадковування тих чи інших ознак від них відхиляється. Основні положення хромосомної теорії спадковості такі:
- гени розташовані в хромосомах у лінійному порядку;
- різні хромосоми мають неоднакові набори генів, тобто кожна з негомологічних хромосом має свій унікальний набір генів;
- кожен ген займає в хромосомі певну ділянку; алельні гени займають у гомологічних хромосомах однакові ділянки;
— усі гени однієї хромосоми утворюють групу зчеплення, завдяки чому деякі ознаки успадковуються зчеплено; сила зчеплення між двома генами, розташованими в одній хромосомі, обернено пропорційна відстані між ними;
— зчеплення між генами однієї групи порушується внаслідок обміну ділянками гомологічних хромосом у профазі першого мейотичного поділу (процес кросинговеру);
— кожен біологічний вид характеризується певним набором хромосом (каріотипом) - кількістю та особливостями будови окремих хромосом.
Як визначається стать тих чи інших організмів під час їхнього індивідуального розвитку - одна з найцікавіших проблем біології.
Як визначається стать різних груп організмів? Ще наприкінці XIX століття вчені звернули увагу на те, що хромосомні набори самців і самок різняться за будовою хромосом однієї з пар. У диплоїдних соматичних (нестатевих) клітинах самок багатьох видів тварин хромосоми всіх пар подібні за будовою, тоді як у самців хромосоми однієї з пар різні. Такі хромосоми, як ви пам'ятаєте, називають статевими хромосомами. Так, у самців дрозофіли одна зі статевих хромосом має паличкоподібну форму (це так звана Х-хромосома), інша - гачкоподібну (Y-xpo-мосома). У самок дрозофіли обидві статеві хромосоми мають однакову будову (Х-хромосоми). Отже, каріотип самок дрозофіли можна умовно позначити як 6А + XX, а самців - 6А + XY (символом «А» позначають нестатеві хромосоми - аутосоми, однакові за будовою в особин різної статі).
Оскільки під час мейозу гомологічні хромосоми розходяться до різних гамет, то у особин однієї статі формується лише один тип гамет (гомогаметна стать), тоді як у особин протилежної - два (гетерогаметна стать). У багатьох групах організмів гомогаметною статтю є жіноча, а гетерогаметною — чоловіча (мухи, клопи, жуки, ссавці, більшість видів риб, деякі земноводні та дводомні рослини тощо), а в інших навпаки (метелики, плазуни, птахи, деякі риби і земноводні). У деяких видів особини різних статей відрізняються за кількістю статевих хромосом. Так, у коників у диплоїдному наборі самки є обидві статеві хромосоми, а самця — лише одна. Отже, у більшості роздільностатевих організмів стать майбутньої особини визначається в момент запліднення і залежить від того, скільки і які зі статевих хромосом поєднуються в зиготі.
Але крім хромосомного, існують й інші механізми Визначення статі організмів. У деяких безхребетних тварин (наприклад, коловерток, багатощетинкового Черв'яка динофілюса) стать майбутньої особини ви значається ще до моменту запліднення. Ці тварини можуть утворювати яйцеклітини двох типів: великі, багаті на жовток, і дрібні, з невеликим запасом поживних речовин. З яйцеклітин першого типу розвиваються лише самки, а з другого - самці.
У певних організмів на формування статі майбутньої особини можуть впливати біологічно активні речовини. Наприклад, у морської червоподібної тварини - бонелії - личинки, які прикріплюються до поверхні дна, розвиваються в великих (до 1 м завдовжки) самок. А ті з них, які потрапляють на - хоботок самки, під впливом її гормонів перетворюються на карликових самців 1-3 мм завдовжки.
У таких суспільних комах, як медоносна бджола, джмелі, мурашки, самки утворюють яйця двох типів: запліднені та незапліднені (партеногенетичні). З яєць першого типу розвиваються самки, а з другого — самці.
У деяких видів риб і земноводних під час зародкового розвитку одночасно закладаються зачатки як чоловічих, так і жіночих статевих залоз. Проте в процесі подальшого розвитку розвивається лише один із цих типів. Подібне явище спостерігається, наприклад, у риб-«чистильників», самці яких мають «гареми» з кількох самок. Після загибелі самця його функції переходять до однієї з самок, в якої з недиферен-ційованих статевих зачатків починають розвиватися сім'яники.
Чим визначається співвідношення статей у популяціях? У популяціях організмів, стать яких визначається в момент запліднення, співвідношення самців і самок, згідно із законом розщеплення, має становити 1:1. 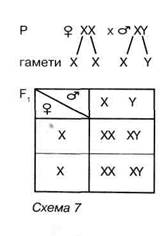
Але в природі таке співвідношення статей рідко спостерігають унаслідок різного рівня смертності самців і самок. Вищу смертність, зазвичай, мають особини гетерогаметної статі, оскільки в Y-хромосомі через її менші розміри немає деяких алельних генів, наявних в Х-хромосомі. Тому у фенотипі особин гетерогаметної статі можуть проявитися летальні або напівлегальні рецесивні алелі. Наприклад, у шовковичного шовкопряда більше самців, ніж самок, оскільки від особливого вірусного захворювання частіше гине гусінь, з якої мали б розвиватися самки (гетерогаметна стать).
Що таке успадкування, зчеплене зі статтю? Існують деякі ознаки, на характер успадкування яких впливає стать організму. Це пояснюється неоднаковим складом генів у X- та Y-хромосом, про що згадували раніше. У Х-хромосомі є ділянки з гемами, яких немає в Y-хромосомі через її менші розміри, хоча в ній можуть бути деякі гени, яких немає в Х-хромосомі (наприклад, ген, який зумовлює наявність або відсутність волосин по краю вушної раковини людини),
У кішок зчеплено зі статтю успадковуються певні види забарвлення шерсті. Відомо, що коти майже ніколи не мають черепахового забарвлення (руді та чорні плями на білому тлі): вони бувають або з темними плямами, або рудими. Це пояснюється тим, що алельні гени, які зумовлюють руде або чорне забарвлення шерсті, розташовані лише в Х-хромосомі. Жодна з алелей не домінує над іншою. Тому кішки, гетерозиготні за цим геном, мають черепахове забарвлення шерсті, на відміну від котів, у яких Y-хромосома його позбавлена.
У людини зчеплено зі статтю успадковується майже 150 ознак, зокрема деякі захворювання (дальтонізм, гемофілія тощо). Дальтонізм (нездатність розпізнавати деякі кольори) визначається рецесивною алеллю, розташованою в X- і відсутньою в Y-хромосомі. Тому чоловік, який має цю алель, хворіє на дальтонізм. У жінок це захворювання виявляється лише в особин, гомозиготних за рецесивною алеллю; гетерозиготні жінки фенотипно здорові, хоча і є носіями цієї алелі.
Так само успадковується і гемофілія (нездатність крові зсідатися, внаслідок чого людина може загинути навіть за незначних ушкоджень кровоносних судин). Як правило, рецесивна алель, яка зумовлює це захворювання, передається з покоління в покоління гетерозиготними жінками-носіями, оскільки гомозиготні за цією алеллю жінки хворіють на гемофілію і не доживають до репродуктивного віку.
Що таке ген? Раніше ви дізналися про хімічну природу гена. Тривалий час, доки не було з'ясовано структуру нуклеїнових кислот і генетичний код, ген вважали неподільною одиницею спадкової інформації, рекомбінацій і мутацій. Але згодом з'ясували, що зміни можуть зачіпати не весь ген, а лише певну його частину. Під час кросинговеру гомологічні хромосоми можуть обмінюватися як цілими генами, так і їхніми частинами. Мінімальна ділянка молекули нуклеїнової кислоти, яка може бути поділена під час кросинговеру, становить усього 1—2 пари нуклеотидів. Проте ген — цілісна функціональна одиниця, оскільки будь-які порушення його структури змінюють закодовану в ньому інформацію або призводять до її втрати.
Серед генів розрізняють структурні, що кодують структуру білків і РНК певних типів, і регуляторні, які слугують місцем приєднання ферментів та інших біологічно активних речовин. Останні впливають на активність структурних генів і беруть участь у процесах подвоєння ДНК і транскрипції. Розміри регуляторних генів порівняно зі структурними, зазвичай, незначні.
Отже, ген — фактор спадковості, функціонально неподільна одиниця генетичного матеріалу у вигляді ділянки молекули нуклеїнової кислоти (ДНК або РНК). Він кодує первинну структуру білка, молекули тРНК чи рРНК або взаємодіє з біологічно активними речовинами (наприклад, ферментами).
Яка організація геному в різних організмів-У різних організмів кількість генів у геномі може значно варіювати. Найпростіше організований гено»1 вірусів. Він може включати від одного гена до кількох сотень генів. Геном прокаріотів містить як структури1' так і регуляторні гени. Наприклад, ДНК кишкової палички складається з 3 800 000 пар нуклеотидів, а кількість структурних генів — приблизно 1 000. Майже половина довжини такої молекули генетичної формації не несе, це ділянки, розташовані між ок-
Мими генами (так звані спейсери; від англ. спейс -простір). Геном еукаріотів складніший, бо має більшу кількість ядерної Днк та структурних регуляторних генів. Так, геном дрозофіли складається з майже 180 000 000 пар нуклеотидів і включає близько 10 000 структурних генів. За даними останніх досліджень, структурних генів у геномі людини близько 30000.
У результаті досліджень геному різних еукаріотич-них організмів з'ясовано, що кількість ДНК в ядрі перевищує необхідну для кодування всіх структурних генів у 8—10 разів. Причини цього явища різні. По-перше, ДНК еукаріотів містить велику кількість послідовностей нуклеотидів, кожна з яких повторюється до сотень тисяч разів. По-друге, значна частина ДНК взагалі не несе генетичної інформації. По-третє, чимало є регуляторних генів, які не кодують структуру білків або РНК.
Що таке цитоплазматична спадковість? У клітинах еукаріотів, крім спадкового матеріалу, розташованого в ядрі, виявлено також цитоплазматичну спадковість, або позаядерну. Вона полягає у здатності певних структур цитоплазми зберігати і передавати нащадкам частину спадкової інформації батьків. Хоча провідна роль в успадкуванні більшості ознак організму належить ядерним генам, роль цитоплазматичної спадковості теж значна. Вона пов'язана з двома видами генетичних явищ:
- успадкуванням ознак, які кодуються позаядерни-ми генами, розташованими в певних органелах (мітохондріях, пластидах);
- проявом у нащадків ознак, зумовлених ядерними генами, але на формування яких впливає цитоплазма яйцеклітини.
Про існування генів в органелах (мітохондріях і Пластидах), здатних до самоподвоєння, стало відомо на початку XX століття під час вивчення зелених і безбарвних пластид у деяких квіткових рослин із мозаїчним забарвленням листків. Позаядерні гени взаємодіють із ядерними і перебувають під контролем ядерної ДНК. Цитоплазматична спадковість,повязана з генами пластид, характерна для різних видів рослин (наприклад, ротиків). Серед таких рослин є форми зі строкатими листками, причому ця ознака передається по материнській лінії. Строкатість листків зумовлена нездатністю частини пластид ут ворювати пігмент хлорофіл. Після поділу клітин із безбарвними пластидами в листках виникають білі плями, які чергуються із зеленими ділянками. Передача строкатості по материнській лінії пояснюється тим, що під час утворення статевих клітин пластиди потрапляють до яйцеклітин, а не до сперміїв. Пластиди, які розмножуються поділом, мають генетичну безперервність: зелені пластиди дають початок зеленим, а безбарвні - безбарвним. Під час поділу клітини пластиди розподіляються випадково, внаслідок чого утворюються клітини з безбарвними, зеленими або обома типами пластид одночасно.
Явище цитоплазматичної спадковості, пов'язаної з генами мітохондрій, вивчали на прикладі дріжджів. У цих мікроорганізмів у мітохондріях виявлено гени, які зумовлюють відсутність або наявність дихальних ферментів, а також стійкість до дії певних антибіотиків.
Вплив ядерних генів материнського організму через цитоплазму яйцеклітини на формування певних станів ознак можна простежити на прикладі ставко-вика. У цього прісноводного молюска є форми з різними станами спадкової ознаки напрямку закрученості черепашки - лівого або правого. Алель, яка визначає правозакрученість черепашки, домінує над алеллю лівозакрученості, але при цьому напрямок закрученості зумовлений генами материнської особини. Наприклад, особини, гомозиготні за рецесивною алеллю лівозакрученості, можуть мати правозакручену черепашку, якщо домінантну алель правозакрученості мав материнський організм.
Який є взаємозв'язок між генами та ознаками, які вони визначають? Тривалий час у генетиці існувало правило, згідно з яким кожний ген визначає синтез одного білка, чи одну ознаку. Проте подальші дослідження показали, що відношення «ген — ознака» значно складніші. Стали відомі явища множинної дії генів і взаємодії неалельних генів.
Раніше ми з вами розглянули різні варіанти взаємодії алельних генів: повне і неповне домінування, проміжний характер успадкування. Проте на формування певних станів ознак часто впливає взаємодія двох або більшої кількості неалельних генів. Така взаємодія можлива у різних формах.
Один із типів взаємодії неалельних генів проявляється в тому, що певна алель одного гена пригнічує прояв алелі іншого, неалельного.
Наприклад, пурпурне забарвлення очей дрозофіли зумовлене рецесивною алеллю одного з генів. Але вона не проявиться у фенотипі, якщо в гомозиготних за цією алеллю особин у гомозиготному стані перебуватиме рецесивна алель іншого неалельного гена, який пригнічуватиме дію першого.
Інший поширений тип взаємодії неалельних генів полягає в тому, що для прояву в фенотипі певного стану ознаки необхідна взаємодія домінантних алелей Двох або більшої кількості неалельних генів.
Так, фіалкове забарвлення плодів баклажана залежить від взаємодії домінантних алелей двох неалельних генів, завдяки чому утворюється відповідний пігмент (антоціан). Якщо хоча б один із цих генів гомозиготний за рецесивною алеллю, пігмент не синтезується і утворюються безбарвні плоди. Подібне явище спостерігається і у запашного горошку, в якого Домінантні алелі двох неалельних генів зумовлюють червоне забарвлення віночка. Але якщо хоча б один із цих генів гомозиготний за рецесивною алеллю, віночок буде білим.
Тварин (наприклад, мишей, кролів) для формування темного забарвлення шерсті потрібні домінантні алелі двох неалельних генів, один із яких вйзначає наявність пігменту, а інший - його розподіл по волосині. Якщо перший з цих генів гомозиготний за рецесивною алеллю, то пігмент не синтезується і народжуються білі особини (альбіноси). У людини розвиток нормального слуху також визначають домінантні алелі двох неалельних генів, один із яких відповідає за нормальний розвиток завитки внутрішнього вуха, а інший - слухового нерва. Якщо хоча б один із цих генів гомозиготний за рецесивною алеллю — людина глуха від народження.
Що таке множинна дія генів? Більшості генів притаманна властивість, коли одна їхня алель впливає на формування певних станів кількох різних ознак. Це явище називають множинною дією генів.
Наприклад, у людини відоме захворювання - арахнодактилія (у перекладі - «павучі пальці»). Воно зумовлене мутацією домінантної алелі, що впливає на формування видовжених пальців на руках і ногах, неправильне положення кришталика ока і вроджені вади серця. У дрозофіли рецесивна алель одного з генів визначає відсутність пігменту очей (білоокість), світле забарвлення тіла, змінює будову статевих органів, знижує плодючість і тривалість життя. У картоплі домінантна алель визначає рожеве забарвлення бульб і червоно-фіалкове - віночка (у рослин, гомозиготних за відповідною рецесивною алеллю, бульби і віночки квіток синюваті або білі).
Генотип особин кожного виду є цілісною системою, хоча і складається з окремих генів, які можуть відокремлюватись один від одного і успадковуватись незалежно. Цілісність генотипу, яка склалася в процесі історичного розвитку виду, проявляється в тому, що формування станів більшості ознак організму є результатом взаємодії алельних і неалельних генів, а алелі більшості генів впливають на розвиток певних станів кількох ознак.
Лекція № 20
Дата добавления: 2022-01-22; просмотров: 48; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!