Биология. В понятие биология человека некоторые учёные включают почти всё содержание антропологии, обогащённое методами и фактами смежных биологических дисциплин.
Скелет человека разумного. Голова большая. На верхних конечностях по пять длинных гибких пальцев, один из которых несколько отстоит от остальных, а на нижних — пять коротких пальцев, помогающих балансировать при ходьбе. Помимо ходьбы, люди также способны к бегу, но, в отличие от большинства приматов, способность к брахиации развита слабо.
Медицинские. Норма. Средний вес мужчины составляет 70—80 кг, женщины — 50—65 кг, хотя встречаются и более крупные люди (некоторые до 500—600 кг)[11]. Средний рост мужчины на планете 175 см. Средний рост женщины — 170 см. Средний рост человека менялся с течением времени. Так, в средние века люди в Европе были ниже ростом, что заметно по размеру рыцарских доспехов того времени.
11 Догоминидные предки человека их морфологические особенности в Эфиопии и позже в Танзании и Кении были обнаружены большие группы гоминид, древность которых определяется в 4—2,8 млн. лет. По этим материалам в 1978 г. был описан новый вид австралопитеков A . afarensis , более примитивный, чем A . africanus , но тоже двуногий и с чертами «промежуточности» по отношению к современным человекообразным обезьянам и человеку (рис. 15.4). Объем мозговой полости черепа оказался в пределах 380—450 см3, т.е. практически не отличался от объема мозга современного шимпанзе.
Действительно, австалопитеки имеют большее сходство с человеком не по объему и строению мозга, а по строению зубной системы и способу передвижения. Изучение австралопитековых показало, что именно двуногость, а не большой объем мозга, явилась ключевой адаптацией ранних гоминид. В 1964 г. по находкам, сделанным в Танзании, был выделен вид Homo habilis , или Человек умелый, имеющий абсолютный возраст 2—1,7 млн. лет. Отличительными чертами его являются двуногость, в целом прогрессивное строение кисти, зубной системы, объем мозговой коробки от 540 до 700 см3, что примерно в полтора раза превышает объем мозга австралопитеков.

Рис. 15.4. Австралопитек афарский. Реконструкция
На внутренней поверхности черепа обнаруживаются признаки прогрессивных нейроморфологических изменений, определяющиеся по отпечаткам головного мозга: выраженная асимметрия полушарий и развитие двух речевых центров как условие для возникновения членораздельной речи. Большой палец стопы не отведен в сторону. Это свидетельствует о том, что морфологические перестройки, связанные с прямохождением, у него полностью завершились. Вместе с остатками Н. habilis найдены орудия труда со следами целенаправленной обработки, свидетельствующие о ранних формах трудовой деятельности.Перечисленные признаки, ведущим из которых является прогрессивное развитие мозга, характеризуют организм уже с иной морфофункциональной организацией по сравнению с австралопитеками. Эти признаки считаются специфичными для рода Homo . Сопоставление морфологии африканского и афарского австралопитеков с Н. habilis и современным человеком позволяет предположить следующую схему филогенетических взаимоотношений ранних гоминид (рис. 15.5). Африканский австралопитек, считавшийся раньше общим предком как рода Homo , так и других австралопитековых, является в этой схеме представителем боковой ветки эволюции, приведшей к узкой специализации и образованию мощных форм типа A . robustus , которые вымерли около 1 млн. лет назад. Общим предком всех австралопитековых и рода Homo является в соответствии с этой схемой A . afarensis . 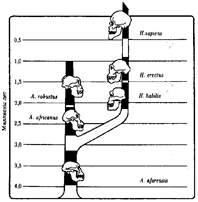
Рис. 15.5. Филогенетические взаимоотношения ранних гомини
Таким образом, на протяжении 1—1,5 млн. лет представители двух близких родов и, возможно, нескольких видов семейства гоминид сосуществовали, причем не только во времени, но и на перекрывающихся территориях. В основе дивергенции различных линий ранних гоминид и австралопитековых могли лежать разного рода механизмы изоляции,
Ведущими факторами эволюции на прегоминидной стадии антропогенеза являлись, несомненно, факторы биологической эволюции, главным из которых является естественный отбор. Об этом свидетельствует, в частности, большое видовое разнообразие австралопитековых, обитавших в различных условиях практически на всей территории Южной, Центральной и Северо-Восточной Африки. В это же время в происхождении рода Homo имело место скачкообразное изменение наследственного материала.
В разных органах и системах прегоминид обнаруживалась асинхронность филогенеза. Есть предположение о том, что эволюция коры больших полушарий мозга состоит из двух компонентов, разобщенных по времени: соматического, обеспечивающего сенсорно-моторные функции, и несоматического, связанного с высшими психическими функциями. Если локомоторный комплекс подвергался длительным постоянным изменениям, то головной мозг эволюционировал скачкообразно. Элементы скачкообразности в эволюции некоторых структур ранних гоминид могли быть обусловлены «транспозиционными взрывами» (перестройками генома за счет подвижных генетических элементов, переносящих комплексы генов на новое место). Это могло сопровождаться постепенным развитием других морфофизиологических свойств в результате накопления малых мутаций под контролем естественного отбора. Но именно в период становления Н. habilis возникла, вероятно, часть хромосомных перестроек в геноме человека, о которых говорилось выше.
12 Следующей ступенью гоминизации после появления Н. habilis считается возникновение архантропов, представителем которых является сборный вид Н. erectus , или Человек прямоходящий. Материальная культура и ярко выраженная социальность позволили ему быстро и эффективно расселиться по всей территории Африки и Евразии и освоить обширный ареал, разнообразный в климатическом отношении. Действительно, орудия труда Н. erectus более прогрессивны, чем у Н. habilis , а масса мозга (от 800 до 1000 г) явно превышает минимальную массу (750 г), при которой возможно существование речи. Наличие при этом речевых центров, возникших впервые у Н. habilis , предполагает и развитие второй сигнальной системы.
Выделяют три группы Н. erectus : обитавшие в Европе, Азии и Африке. Долгое время древнейшими архантропами считались азиатские представители из Индонезии и Восточного Китая — питекантроп и синантроп. Однако находки последних лет на территории Израиля (1982) и Кении (1984), датирующиеся соответственно 2,0 и 1,6 млн. лет, сопровождающиеся элементами материальной культуры и признаками использования огня, показали, что арогенная эволюция гоминид происходила на африканском континенте и на Ближнем Востоке. Это позволило связать происхождение Н. erectus с восточноафриканскими формами Н. habilis .
В связи с тем что небольшое различие ископаемых остатков Н. erectus не соответствует значительному разнообразию природно-климатических условий указанных территорий, можно заключить, что в этих адаптациях значительную роль играли наряду с факторами биологической эволюции также и социальные факторы: совместное изготовление укрытий, орудий труда и использование огня.
13 Роль Н. erectus как этапа антропогенеза никогда не подвергалась сомнению. Что же касается палеоантропа, или неандертальского человека, то его роль в происхождении человека в настоящее время оспаривается. Это связано в первую очередь с обнаружением большого количества ископаемых остатков человека с промежуточными чертами между Н. erectus и человеком современного физического типа. Кроме того, палеонтологические находки последних лет дают повод судить о недооценке интеллектуальных возможностей неандертальцев. На всех стоянках обнаружены следы костров и обгоревшие кости животных, что свидетельствует об использовании огня для приготовления пищи. Орудия труда их гораздо совершеннее, чем у предковых форм. Масса мозга неандертальцев около 1500 г, причем сильное развитие получили отделы, связанные с логическим мышлением. Костные остатки неандертальца из Сен-Сезер (Франция) были найдены вместе с орудиями труда, свойственными верхнепалеолитическому человеку, что свидетельствует об отсутствии резкой интеллектуальной грани между неандертальцем и современным человеком. Имеются данные о ритуальных захоронениях неандертальцев на территории Ближнего Востока.
Эти и ряд других находок позволили в конце 60-х годов выделить палеоантропов в отдельный подвид Homo sapiens neanderthalensis в отличие от неоантропа H . s . sapiens , который, таким образом, тоже получил ранг подвида. Наиболее древние ископаемые остатки его возрастом 100 тыс. лет обнаружены также на территории Северо-Восточной Африки. Многочисленные находки палеоантропов и неоантропов на территории Европы, датирующиеся 37—25 тыс. лет, свидетельствуют о существовании обоих подвидов в течение нескольких тысячелетий.
H.s. neanderthalensis в виде ископаемых остатков не обнаруживается позже рубежа в 25 тыс. лет. Быстрое исчезновение палеоантропов может быть объяснено вытеснением их людьми с более совершенной техникой изготовления орудий труда и метисацией с ними.
С возникновением человека современного физического типа роль биологических факторов в его эволюции свелась к минимуму, уступив место социальной эволюции. Об этом отчетливо свидетельствует отсутствие существенных различий между ископаемым человеком, жившим 30—25 тыс. лет назад, и нашим современником.
1 приматы – размер с кошку, длинный хвост, конечности с когтями, мордочка удлинённая. 40-50 млн. лет назад примитивные приматы дали начало обезьянам нового и старого света (широконосые и узконосые).
Парапиты- проприонты (древесный образ жизни, размер тела с собаку, похожи на гиббонов).
Дриопитекум (древесная обезьяна) имеет большой размер тела и большой вес.
Рамопитеки. Прямохождение, развитая ягодичная мускулатура, мощность коленного сустава.
Австралопитеки. 50кг. рост 150см. прямоходящие, масса мозга – 500г., всеядные.
Гомо гапиес – масса мозка – 700г. умеют изготавливать орудия труда.
Гомо эректус (человек прямоходящий) – низкий покатый лоб, масса мозга – 800г. рост – 150-160см. трудовая деятельность.
Гомо сапиенс (человек разумный) возник около
130 лет назад.
14 понятие о расах и видовое единство человечества
Человечество в настоящее время представлено одним видом Homo sapiens (Человек разумный). Однако этот вид неоднороден. Он полиморфный и состоит из трех больших и множества мелких переходных рас — биологических групп, отличающихся мелкими морфологическими признаками. К таким признакам относятся: тип и цвет волос, цвет кожи, глаз, форма носа, губ, лица и головы, пропорции тела и конечностей.
Расы появились в результате расселения и географической изоляции предков современных людей в разных природно-климатических условиях. Расовые признаки наследственны, Они возникли в далеком прошлом под непосредственным влиянием среды и носили адаптивный характер. Выделяют следующие большие расы.
Негроидная (австрало-негроидная, или экваториальная) раса характеризуется темным цветом кожи, курчавыми или волнистыми волосами, широким и мало выступающим носом, толстыми губами и темными глазами. До эпохи колонизации эта раса была распространена в Африке, Австралии и на островах Тихого океана.
Европеоидная (евро-азиатская) раса отличается светлой или смуглой кожей, прямыми или волнистыми волосами, хорошим развитием волосяного покрова на лице у мужчин (борода и усы), узким выступающим носом, тонкими губами. Представители этойрасы расселены в Европе, Северной Африке, Передней Азии и Северной Индии.
Для монголоидной (азиатско-американской) расы характерны смуглая или светлая кожа, прямые, часто жесткие волосы, уплощенное широкое лицо с сильно выступающими скулами, средняя ширина губ и носа, заметное развитие эпикантуса. Первоначально эта раса заселяла Юго-Восточную, Восточную, Северную и Центральную Азию, Северную и Южную Америку.
Хотя большие расы заметно отличаются друг от друга по комплексу внешних признаков, они связаны между собой рядом промежуточных типов, незаметно переходящих один в другой.
Как уже отмечалось, расовые особенности наследственны, и, по-видимому, часть из них в прошлом носила адаптивный характер. Так, темная кожа негроидов предохраняет организм от ярких солнечных лучей; в курчавых волосах создаются воздушные прослойки, защищающие от жары. Широкий нос и толстые вздутые губы с большой поверхностью слизистых оболочек способствуют быстрому испарению влаги с высокой теплоотдачей. Светлая кожа европеоидов пропускает ультрафиолетовые лучи и этим способствует синтезу витамина D, предохраняя человека от рахита. Узкий выступающий нос обеспечивает согревание вдыхаемого воздуха. Некоторые признаки монголоидов являются результатом адаптации к суровому, часто с пылевыми бурями климату Центральной Азии.
О биологическим единстве человеческих рас свидетельствуют:
1. отсутствие генетической изоляции и неограниченные возможности скрещиваний с образованием плодовитого потомства;
2. равноценность рас в биологическом и психологическом отношении;
3. наличие переходных рас между большими расами, совмещающими признаки двух соседних.
4. локализация на втором пальце кожных узоров типа дуг (у человекообразных обезьян — на пятом); у всех представителей рас одинаковый характер расположения волос на голове и другие морфофизиологические признаки.
15 современная классификация,распространение и морфологические особенности человеческих рас
Морфологические и в меньшей степени физиологические признаки дают возможность выделить внутри человечества три основные большие расы: европеидную, австрало-негроидную и монголоидную.
Европеоиды имеют светлую или смуглую кожу, прямые или волнистые волосы, узкий выступающий нос, тонкие губы и развитый волосяной покров на лице и теле. У монголоидов кожа также может быть как светлой, так и темной, волосы обычно прямые, жесткие, темно пигментированные, косой разрез глаз и эпикант («третье веко»). Негроиды характеризуются темной кожей, курчавыми или волнистыми волосами, толстыми губами и широким, слегка выступающим носом. Имеются отличия рас и по некоторым физиологическим и биохимическим показателям: интенсивность потоотделения с единицы площади кожи у негроидов выше, чем у европеоидов, средние показатели уровня холестерина в плазме крови наиболее велики у европеоидов.
В рамках каждой большой расы выделяются отдельные антропологические типы с устойчивыми комплексами признаков, называющиеся малыми расами. Существует три основных подхода к классификации рас: без учета их происхождения, с учетом происхождения и родства и на основе популяционной концепции. В соответствии с первым подходом три большие расы включают в себя 22 малые, причем между большими расами располагаются по две переходные малые. Схема расовой классификации изображается при этом в виде круга (рис. 15.7). Несмотря на то что при такой классификации не учитывается происхождение рас, само существование малых переходных рас, сочетающих в себе одновременно признаки двух больших рас (эфиопская, южносибирская, уральская и т.д.), свидетельствует, с одной стороны, о динамизме расовых комплексов признаков, а с другой — об условности членения человечества даже на большие расы.
Классификация с учетом происхождения рас изображается в виде эволюционного древа с коротким общим стволом и расходящимися от него ветвями (рис. 15.8). В основе таких классификаций лежит обнаружение черт архаизма и эволюционной продвинутости отдельных рас, в соответствии с чем разные большие и малые расы занимают разное положение на ветвях такого древа. Выявление архаичных и прогрессивных черт среди морфологических признаков носит субъективный характер, благодаря чему схемы расовых классификаций такого рода очень многообразны. Но самым большим недостатком подхода к классификации рас исходя из их происхождения является попытка расположить расы на разных уровнях эволюционного древа, т.е. признание их биологической неравноценности.
Кроме того, данные палеоантропологических исследований показывают, что вплоть до верхнего палеолита на территориях, обитаемых людьми, практически нигде не сформировались расовые типы человека, с которыми были бы генетически связаны современные большие расы. Это подтверждает анализ верхнепалеолитических находок скелетов людей современного физического типа из сунгирских погребений (Россия), живших приблизительно 26 тыс. лет назад. Все черепа, принадлежащие им, характеризуются мозаичным сочетанием расовых признаков и не могут быть отнесены ни к одной из современных рас.
что процесс формирования расовых признаков — довольно поздний, идущий параллельно в разных регионах на рубеже верхнего палеолита — мезолита на фоне исходной разнородности расовых признаков у человека современного физического типа.
Первичное появление на протяжении эволюции признаков малых, а не больших рас позволяет сделать вывод о том, что европеоидная, монголоидная и негроидная расы имеют мозаичное происхождение и представляют собой крупные популяции, объединенные не столько общностью происхождения, сколько климатогеографическими характеристиками условий существования и адаптивностью большинства основных признаков.
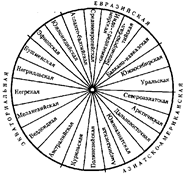
Рис. 15.7. Расовая классификация человечества без учета происхождения рас
Это заключение хорошо согласуется с популяционной концепцией рас. Суть ее заключается в следующем. Если принять, что большие расы человека представляют собой огромные популяции, то малые расы — субпопуляции больших, локальные естественные общности людей внутри которых — конкретные этнические образования (нации, народности) — являются более малыми популяциями. Если предположить при этом, намеренно упрощая ситуацию, что этносы не разделяются на элементарные популяции, и считать их просто состоящими из конкретных особей, то получится сложная структура, включающая в себя четыре уровня иерархии (рис. 15.9).
Рис. 15.9. Расы как выражение генетического полиморфизма человечества:
1—отдельные индивидуумы, 2—этносы, 3—малые расы, 4—большие расы
На основании исследований распределения различных групп крови и белков в популяциях человека произведено сравнение доли каждого из четырех уровней меж- и внутрипопуляционных различий в общем объеме генетического полиморфизма человека по этим признакам.
| Уровни различий | Доля генетического разнообразия, % |
| Индивидуумы Этносы Малые расы Большие расы Итого | 84 5 3 8 100 |
Таким образом, от тотального генетического полиморфизма человечества расовые признаки составляют только 8%, в то время как основная доля генетического разнообразия определяется многообразием отдельных индивидуумов. Иными словами, немец может быть генетически гораздо ближе к полинезийцу, чем к другому немцу, живущему в соседней квартире.
биохимический полиморфизм человека эволюционно возник раньше и развивался дольше по сравнению с возникновением комплексов расовых признаков. Из этого следует, что расы не представляют собой особых изолированных групп людей, характеризующихся наборами специфических генов. Расовые же характеристики являются не более чем отдельными проявлениями общего генетического полиморфизма, выражающегося в первую очередь в сложных морфологических признаках. Некоторые из них адаптивны, другие сформировались на основе коррелятивной изменчивости, но все они касаются лишь ряда второстепенных особенностей (цвета кожи, волос, глаз и т.д.) и не затрагивают таких общечеловеческих признаков, как морфология головного мозга, а также строение и функции руки как органа труда.
На основании определения числа аллелей, свойственных той или иной группе организмов, возможно определение генетического расстояния между ними. Эта величина для больших рас человека составляет 0,03. Она гораздо ниже цифр, характерных для истинных подвидов (0,17—0,22), и еще более мала по сравнению с межвидовым расстоянием (0,5—0,6 и более). В животном мире генетическое расстояние, равное 0,03, соответствует обычно генетическим отличиям местных популяций друг от друга. Все эти данные свидетельствуют о том, что понятие расы условно, второстепенно и не позволяет подводить под иерархическую классификацию рас глубокую биологическую, а значит, и социальную базу.
16 Социал-дарвинизм и расизм,как антинаучные концепции
РАСИЗМ
(определение из Большой Советской энциклопедии)
Расизм - совокупность антинаучных концепций, основу которых составляют положения:
[1] о физической и психической неравноценности человеческих рас и
[2] о решающем влиянии расовых различий на историю и культуру человеческого общества.
Для всех разновидностей расизма характерны ложные человеконенавистнические идеи об исконном разделении людей на высшие и низшие расы, из которых первые якобы являются единственными создателями цивилизации, призванными к господству, а вторые, напротив, не способны к созданию и даже усвоению высокой культуры и обречены быть объектами эксплуатации.
Представления о природном неравенстве рас возникли в рабовладельческом обществе, где они служили для обоснования социальных различий между рабовладельцами и рабами. В средние века утверждения о «кровных» различиях между «знатью» и «чернью» призваны были оправдать сословное неравенство. В эпоху первоначального накопления капитала (16-18 вв.), когда европейские государства впервые захватили колонии, Р. служил целям бесчеловечной эксплуатации, а часто и обоснованием истребления индейцев Америки, африканцев, многих народов Южной Азии, Австралии и Океании.
В середине 19 в. появились первые обобщающие «труды» по Р. В сочинении «Опыт о неравенстве человеческих рас» Ж. А. Гобино объявил «высшей» расой светловолосых и голубоглазых арийцев, которых он считал создателями всех высоких цивилизаций, сохранившимися в наиболее «чистом» виде среди аристократии германских народов. Теория Гобино, основанная на неправомерном отождествлении рас и языковых семей, стала краеугольным камнем многих расистских концепций.
В США Д. Нотт и Д. Глиддон в книге «Типы человечества» (1865) с позиции полигенизма пытались доказать расовую неполноценность негров
В дальнейшем расистские идеи тесно переплелись с социальным дарвинизмом, представители которого переносили учение Ч. Дарвина о естественном отборе и борьбе за существование на человеческое общество (Д. Хайкрафт и Б. Кидд в Великобритании, Ж. Лапуж во Франции, Л. Вольтман, Х. Чемберлен иО. Аммон в Германии, М. Грант в США и др.).
В своих построениях социал-дарвинисты широко использовали мальтузианство, а также положения евгеники для обоснования превосходства наследственных свойств господствующих классов по сравнению с трудящимися, полезности искусственного подбора брачных пар для улучшения расы и необходимости принудительной стерилизации лиц, которые будут признаны «генетически неполноценными». Все эти человеконенавистнические утверждения получили особенно широкое распространение в эпоху империализма в Великобритании, Германии и США.
После 1-й мировой войны 1914-18 главным образом в Германии в реакционных кругах приобрёл популярность «нордический миф» о превосходстве над всеми др. расами северной или нордийской, расы (см. Нордизм) высокорослых длинноголовых блондинов, якобы связанной генетически с народами, говорящими на германских языках. Этот вариант Р., восходящий к концепциям Гобино и Лапужа, пропагандировался в сочинениях многих немецких лжеучёных, откровенно поддержавших нацизм (Х. Гюнтер и др.). Однако ввиду несоответствия внешности большинства нацистов (начиная с Гитлера) нордийскому расовому типу идеологи немецкого фашизма всё чаще стали говорить не о длинноголовых высокорослых блондинах, а о «северной расовой душе» или просто «высшей расе», в которую включались также итальянские фашисты и японские милитаристы.
В годы гитлеровской диктатуры в Германии Р., ставший официальной идеологией фашизма, использовался для оправдания захвата чужих земель, физического уничтожения многих миллионов мирного населения (в первую очередь в СССР и слав. странах), заключения в концлагеря, пыток и казней антифашистов в самой Германии. Подобная же «расистская практика» проводилась японскими милитаристами в Китае и др. азиатских странах, итальянскими фашистами в Эфиопии, Албании, Греции.
Разгром фашизма нанёс Р. и его кровавой практике сокрушительный удар, в полной мере вскрыл человеконенавистническую сущность мифа о «высшей германской расе».
После 2-й мировой войны 1939-45 сторонники Р. пытаются использовать тесты умственных способностей (см. Коэффициент интеллектуальности) разных расовых групп с целью «доказать» их психическую неравноценность (американские психологи Г. Гарретт, О. Шай, Н. Дженсон и др.). В действительности результаты тестов предопределяются не расовой принадлежностью исследуемых, но их социальным положением, условиями труда и быта (например, работа О. Клайнеберга «Раса и психология», 1956).
Р. используется в современных капиталистических странах (особенно в ЮАР, Родезии) для оправдания расовой дискриминации, сегрегации, а во многих случаях и геноцида.
В США усиливается борьба с теорией и практикой Р. как негров и индейцев, так и прогрессивных слоев белых американцев.
В ЮАР и Родезии существуют законы и активно осуществляются запретительные и репрессивные меры, препятствующие общению и заключению браков между белыми, с одной стороны, чёрными и цветными - с другой. Политика сегрегации и апартеида, проводимая в этих странах, сопровождается кровавыми инцидентами, убийствами, жестокими преследованиями противников Р.
[!] Из 22 млн. чел. населения ЮАР (1970) только белые (3,8 млн., около 20% населения) являются полноправными гражданами, остальные же - африканцы, «цветные» (метисы) и выходцы из Азии (главным образом индийцы) - подвергаются юридически закрепленной дискриминации во всех сферах труда и быта. Аналогичное положение характерно и для Родезии, где Р. объявлен официальной идеологией, призванной увековечить господство европейского меньшинства (менее 300 тыс. чел.) над африканским большинством (свыше 4 млн. чел.).
Во многих других странах различные разновидности Р. используются для обоснования всевозможных националистических концепций, разжигания вражды между народами. До краха фашистской диктатуры в Португалии (1974) расовой дискриминации подвергалось коренное африканское население Анголы, Мозамбика и Гвинеи-Бисау.
[!!!] Антинаучный миф о существовании единой мировой еврейской нации взят на вооружение сионизмом.
В Канаде расовой дискриминации подвергаются эскимосы, индейцы и выходцы из Африки, в Австралии - сохранившиеся в этой части света (около 80 тыс. чел.) аборигены. В Великобритании после 2-й мировой войны 1939-45 усилилась дискриминация «цветных» иммигрантов из стран Британского содружества, число которых в 1974 составляло свыше 1,5 млн. чел. Смыкаясь с идеологами империализма, маоисты для обоснования своей великодержавной политики пропагандируют теорию о расовом единстве народов Азии и исключительности Китая, якобы призванного быть гегемоном мировой истории.
Общественно-политическая и научная борьба со всеми разновидностями Р. развернулась, как только стали появляться расистские сочинения. Активное участие в ней приняли передовые общественные деятели, писатели и учёные всех стран. Фактические материалы, накопленные различными общественными и естественными науками, антропологией, этнографией и др. дисциплинами, изучающими расы и народы, показали полную несостоятельность Р. Все морфологические и физиологические признаки, по которым выделяются расы, малосущественны для общей биологической эволюции и исторического развития человечества.
Исторический опыт свидетельствует, что социально-экономический и культурный прогресс обусловлен не расовым составом населения, а социальным строем.
Практика строительства социализма и коммунизма в СССР, народы которого расово крайне неоднородны, доказывает полную необоснованность концепции о существовании «высших» и «низших» рас и о расовой обусловленности исторического процесса.
Против Р. свидетельствуют также экономические и культурные успехи др. социалистических государств Европы, ДРВ, КНДР, Кубы, развивающихся стран Азии, Африки и Латинской Америки, население которых принадлежит к разным расам и всевозможным смешанным и переходным группам между ними.
[!!!] По решению 21-й сессии Генеральной Ассамблеи ООН в память жертв расстрела в 1960 в Шарпевиле и др. городах ЮАР мирных демонстраций африканцев ежегодно 21 марта проводится международный день борьбы за ликвидацию расовой дискриминации.
Созванные ЮНЕСКО совещания экспертов-специалистов в Москве (1964) и Париже (1967) осудили все виды Р. 25-я сессия Генеральной Ассамблеи ООН приняла резолюцию, в которой подтверждает «твердую решимость добиться полной ликвидации расовой дискриминации и расизма, против которых восстают совесть и чувство справедливости всего человечества» [Документ A (RLS) 2646 (XXV), 4, ХП, 1970]. 1971 был объявлен Международным годом согласованных действий против всех видов расовой дискриминации, а 1974-84 - десятилетием максимально эффективной борьбы с Р.
Дата добавления: 2022-01-22; просмотров: 117; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!