Второй закон термодинамики в открытых системах
Оглавление
Раздел 2. Признаки живого. Обмен, регуляция, адаптация. 10
2.1. Гомеостаз, обмен, негэнтропия. 10
2.1.1. Первый закон термодинамики в открытых системах. 10
2.1.2. Второй закон термодинамики в открытых системах. 15
2.1.3. Стационарное состояние. 16
2.1.4. Работоспособность биологических, систем. 18
2.2. Саморегуляция. 19
Вопросы
| На «3» | + на «4» | + на «5» | |
| 1. | Происходит ли движение и превращение энергии в биологических системах? | А в небиологических? | Существуют ли принципиальные отличия обмена энергией и веществом у живых и неживых систем? |
| 2. | 1 закон термодинамики | Какую энергию могут усваивать автотрофы и гетеротрофы? | Человек за сутки получил с пищей 2000 ккал, а на жизнеобеспечение потратил 1600 ккал. Как изменилась его внутренняя энергия? |
| 3. | Почему «солнцееды» долго не живут? | Почему растения в темноте жить не могут, а грибы – могут? | Почему автотрофы в пищевых цепочках всегда внизу? |
| 4. | Что такое анаболизм и катаболизм? | Существуют ли живые системы, у которых все реакции – анаболические или все – катаболические? | В каком возрасте преобладает анаболизм, а в каком – катаболизм? |
| 5. | 2 основные причины алиментарных дисбалансов. | Какой из процессов – ана- или катаболизм усиливается при лихорадке? | Правда ли, что первичная теплота – это основной обмен, а вторичная – это регулируемый обмен? |
| 6. | Чем различаются термодинамическое равновесие и стационарное состояние? | Может ли живая система перейти в состояние термодинамического равновесия? | Может ли живая система никогда не переходить в состояние термодинамического равновесия? |
| 7. | 2 закон термодинамики | Представьте систему с КПД = 100%. Будет ли она выделять тепло? | Зачем живым существам градиенты? |
| 8. | Что такое энтропия? | Может ли человек создать систему с отрицательной энтропией? | Подчиняются ли живые существа 2 началу термодинамики? |
| 9. | Принцип Пригожина и принцип Ле-Шателье. | Что такое автостабилизация? | Что такое информация? |
| 10. | Что такое гомеостаз? | Различия между гуморальным и нервным механизмами регуляции. | Уровни нейрогуморальной регуляции. |
| 11. | Что такое положительная и отрицательная БОС? | Типы регуляции. | Недостатки регуляции по отклонению и по возмущению. |
Раздел 2. Признаки живого. Обмен, регуляция
Обмен, негэнтропия
Одна из 2 важнейших характеристик живых существ – их тесная связь с окружающей средой, без которой они не могут существовать. Связь эта проявляется в постоянном двустороннем обмене веществом и Е.
Первый закон термодинамики в открытых системах
С точки зрения термодинамики[1], биообъекты – это гетерогенные открытые[2] неравнвесные системы[3]. Обмен вещества и энергии в этих системах происходит не хаотично и случайно, а в соответствии с фундаментальными законами термодинамики. Хотя они и состоят из молекул, но не являются их простой суммой, а обладают новыми свойствами, не характерными для их составляющих элементов. Характеристиками молекул являются их масса, скорость, кинетическая энергия, потенциалы взаимодействий. На макроскопическом уровне эти характеристики отражаются в нагревании, плавлении, диффузии и т.д. Одна из важных макроскопических характеристик системы – ее внутренняя энергия (кинетическая энергия хаотического движения молекул, потенциальная энергия их взаимодействия и внутримолекулярная энергия). Количество энергии, переданное одним телом другому, определяется работой, которую тела совершали друг над другом, или количеством переданной теплоты при неизменных внешних параметрах. Оба способа могут привести к изменению внутренней энергии системы.
 1 закон термодинамики – закон сохранения материи и энергии. Внутренняя энергия системы вместе с ее окружением остается постоянной, но она может перемещаться от одной части системы к другой или переходить из одной формы в другую. Иными словами, Е не возникает и не исчезает, а только виды Е превращаются один в другой (см. таблицу). Иными словами, в открытых системах изменение внутренней энергии равно разности между полученным количеством теплоты Q и работой A, совершаемой самой системой: ∆U=Q-A. Иными словами, количество теплоты, полученное системой, идёт на изменение её внутренней энергии и совершение работы против внешних сил Q = ∆U + A.
1 закон термодинамики – закон сохранения материи и энергии. Внутренняя энергия системы вместе с ее окружением остается постоянной, но она может перемещаться от одной части системы к другой или переходить из одной формы в другую. Иными словами, Е не возникает и не исчезает, а только виды Е превращаются один в другой (см. таблицу). Иными словами, в открытых системах изменение внутренней энергии равно разности между полученным количеством теплоты Q и работой A, совершаемой самой системой: ∆U=Q-A. Иными словами, количество теплоты, полученное системой, идёт на изменение её внутренней энергии и совершение работы против внешних сил Q = ∆U + A.
Живые организмы, как и физические открытые системы, постоянно преобразуют энергию. Фототрофы (растения и некоторые бактерии) способны напрямую утилизировать солнечную энергию и запасать ее, преобразуя в химическую в процессе фотосинтеза. Например 6СО2 + 6Н2О = С6Н12О6. Основным накопителем энергии у них служит крахмал.
Хемотрофы (бактерии) получают Е при окислении неорганических веществ. Например, железобактерии окисляют двухвалентное железо до трёхвалентного: 4Fe(HCO3)2 + O2 + 2H2O = 4Fe(OH)3 + 8CO2. Поскольку энергии выделяется немного, железобактериям приходится окислять большое количество железа, которое со временем образует залежи на дне болот и морей. Помимо них, к хемотрофам относятся серобактерии, нитрифицирующие бактерии, водородобактерии. Растения и бактерии, синтезирующие органические вещ-ва из неорганических, называют автотрофами(«сам себе пища»J). Они же обычно являются продуцентами биомассы и занимают базовое положение в пищевых цепочках.

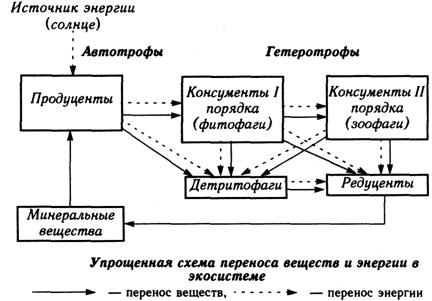 Гетеротрофы(животные, грибы, бактерии, некоторые растения) могут усваивать только химическую энергию, поэтому вынуждены постоянно питаться растениями или другими животными (а иногда и продуктами их жизнедеятельности L, как сапротрофы). Консументы преобразуют одни молекулы в другие в процессе окислительно-восстановительных реакций. Редуценты разрушают органику до минеральных веществ. Пища, окисляясь в организме, дает столько же энергии, как и при сгорании, что доказывает физико-химическую природу биообмена. Основными энергонесущими нутриентами являются белки, жиры и углеводы. При диссимиляции 1 г белка или 1 г углеводов организм аккумулирует 4 ккал энергии (1ккал = 4,18 кДж). Жиры имеют более существенный энергетический потенциал – распад 1 г жира соответствует 9 ккал. В наибольшей степени организм использует с энергетическими целями углеводы и жиры. При выраженном дефиците этих двух макронутриентов в качестве источника энергии кратковременно может быть использован белок пищи. В организме животных энергия запасется главным образом в виде жира (различные депо) и белка (в первую очередь в виде мышечной массы). Запасы углеводов практически отсутствуют (за исключением небольшого количества гликогена).
Гетеротрофы(животные, грибы, бактерии, некоторые растения) могут усваивать только химическую энергию, поэтому вынуждены постоянно питаться растениями или другими животными (а иногда и продуктами их жизнедеятельности L, как сапротрофы). Консументы преобразуют одни молекулы в другие в процессе окислительно-восстановительных реакций. Редуценты разрушают органику до минеральных веществ. Пища, окисляясь в организме, дает столько же энергии, как и при сгорании, что доказывает физико-химическую природу биообмена. Основными энергонесущими нутриентами являются белки, жиры и углеводы. При диссимиляции 1 г белка или 1 г углеводов организм аккумулирует 4 ккал энергии (1ккал = 4,18 кДж). Жиры имеют более существенный энергетический потенциал – распад 1 г жира соответствует 9 ккал. В наибольшей степени организм использует с энергетическими целями углеводы и жиры. При выраженном дефиците этих двух макронутриентов в качестве источника энергии кратковременно может быть использован белок пищи. В организме животных энергия запасется главным образом в виде жира (различные депо) и белка (в первую очередь в виде мышечной массы). Запасы углеводов практически отсутствуют (за исключением небольшого количества гликогена).
Все реакции, протекающие в организме, можно разделить на 2 разновидности. Эндергонические реакции требуют притока энергии и лежат в основе анаболизма. Анаболизм (пластический обмен, ассимиляция) – совокупность реакций синтеза сложных веществ из более простых (синтез глюкозы, АТФ, реакции матричного синтеза). Наиболее важным метаболическим процессом пластического обмена является фотосинтез.
В экзергонических реакциях (расщепление глюкозы и АТФ) энергия выделяется и может использоваться в других реакциях или процессах (в эндергонических). Такие реакции протекают самопроизвольно и часто бывают необратимыми, поскольку часть энергии преобразуется в тепло и рассеивается. В организме совокупность экзергонических реакций называют катаболизмом. Энергия, высвобождающаяся при распаде органических веществ, не сразу используется клеткой, а запасается в форме высокоэнергетических соединений, как правило, в форме АТФ. Синтез АТФ происходит в клетках всех организмов в процессе фосфорилирования, т.е. присоединения неорганического фосфата к АДФ. Катаболизм делится на несколько этапов: 1) подготовительный этап (расщепление сложных углеводов до глюкозы, жиров до жирных кислот и глицерина, белков до аминокислот); 2) бескислородный этап дыхания – гликолиз, в результате глюкоза расщепляется до ПВК (пировиноградной кислоты); в итоге образуется 2АТФ. 3) кислородный этап – дыхание – полное окисление ПВК осуществляется в митохондриях эукариот в присутствии кислорода и включает две стадии: цепь последовательных реакций – цикл Кребса (цикл трикарбоновых кислот) и цикл переноса электронов; в итоге образуется 36АТФ.

Анаболизм и катаболизм неразрывно связаны. Например, фосфорилирования глюкозы свободным фосфатом с образованием глюкозо-6-фосфата является эндергонической реакцией: Глюкоза + Н3РО4 + 13,8 кДж\моль → Глюкозо-6-фосфат + Н2О. 13,8 кДж необходимо затратить, чтобы присоединить фосфатный остаток к глюкозе. Самопроизвольно такая реакция осуществиться не может, необходимо её сопряжение с другой реакцией, в которой энергия выделяется, например, с гидролизом АТФ: АТФ → АДФ + Н3РО4 + 30,5 кДж\моль. При сопряжении этих двух реакций, фосфорилирование глюкозы легко протекает в физиологических условиях: Глюкоза + АТФ → Глюкозо-6-фосфат + АДФ + 30,5 кДж\моль. При этом из 30,5 кДж, выделившихся при расщеплении АТФ, для синтеза Глюкозо-6-фосфата использутся только 13,8 кДж. Остальная энергия рассеивается в виде тепла (первичная теплота). Из-за этого равновесие реакции сильно сдвинуто вправо и она практически необратима. Большинство химических реакций в  открытой системе являются необратимыми, поскольку их конечные продукты (в т.ч. и Е) переходят в недоступное состояние (удаляются в окружающую среду или выпадают в осадок). [studopedia.ru].Т.о., в живых системах эндергонические реакции текут за счет энергии, освобождающейся в экзергонических реакциях. Анаболические (биосинтетические) процессы, требующие притока энергии, текут за счет энергии катаболических процессов распада молекул. При превращении энергии происходит образование тепла.
открытой системе являются необратимыми, поскольку их конечные продукты (в т.ч. и Е) переходят в недоступное состояние (удаляются в окружающую среду или выпадают в осадок). [studopedia.ru].Т.о., в живых системах эндергонические реакции текут за счет энергии, освобождающейся в экзергонических реакциях. Анаболические (биосинтетические) процессы, требующие притока энергии, текут за счет энергии катаболических процессов распада молекул. При превращении энергии происходит образование тепла.
Различают два вида теплоты. Первичная – результат теплового рассеивания энергии биохимических реакций. Вторичная – результат теплового рассеивания энергии при совершении разных форм работы: поддержание осмотичности, свечение, совершение механической работы, а особенно химический термогенез (в буром жире у животных, впадающих в зимнюю спячку, и грудных детей). Вторичная теплота пропорциональна активности ткани, причем основным источником вторичной теплоты будут являться мышцы (поэтому мышечный термогенез мы испльзуем, когда замерзаем). Какую бы работу мы не совершали, конечный результат – тепловая энергия. Поэтому количество энергии, поступающей с пищей, и количество выделяемого тепла примерно одинаковы (Лавуазье и Лаплас, 1784)[4].
Соотношение между поступающей с пищей энергией и её затратами называют энергетическим балансом. Показателями нормального энергетического баланса у детей являются оптимальные показатели роста и развития, а у взрослых – стабильность массы тела.
Затраты энергии принято делить на нерегулируемые: основной обмен и специфическое динамическое действие пищи (пищевой термогенез), и регулируемые: расход энергии на умственную и физическую деятельность. Основой обмен – это количество энергии, затрачиваемой организмом на осуществление основных, непрерывно идущих в нем жизненных процессов (клеточного метаболизма, дыхания, кровообращения, внутренней и внешней секреции, нервной проводимости, мышечного тонуса) в состоянии покоя (например сна). ОО зависит от пола, возраста, веса тела и роста, конституции, гормонального фона. На ОО оказывает влияние время суток, время года и климат. У мужчин он на 10% выше, чем у женщин, у лиц молодого возраста выше, чем у пожилых. У лиц с возбудимой нервной системой, а также у астеников основной обмен усилен, в отличие от лиц с преобладанием тормозных процессов в нервной системе и гиперстеников. В среднем основной обмен в сутки у взрослого человека составляет у мужчин 1 ккал/ час на 1 кг веса, у женщин – 0,9 ккал/час на 1 кг.
При патологи могут наблюдаться значительные отклонения от нормальных величин в ту или иную сторону, что имеет большое диагностическое значение. При заболеваниях щитовидной железы, туберкулезе, острых инфекциях, лихорадочных состояниях, ОО значительно увеличен. При базедовой болезни основной обмен может возрастать вдвое, чем объясняется значительное похудание больных, несмотря на повышенный аппетит. При пониженной функции щитовидной железы, климаксе, ожирении основной обмен снижается. Ярким примером может служить часто наблюдаемое в клинике «климактерическое ожирение».
К регулируемым энергозатратам относится расход энергии на умственную и физическую деятельность (УФД). Затраты энергии тем выше, чем интенсивнее труд. При спокойном сидении обмен веществ увеличивается на 12%, при стоянии – на 20%, при спокойной ходьбе – на 100%, при беге – на 400%. Определяя общий калораж пищи, надо руководствоваться величиной основного обмена веществ, дополнительного обмена веществ и специфического динамического действия принятой пищи.
При дефиците поступающей с пищей энергии со временем (в течение недель, месяцев) развивается дефицит массы тела за счет потерь жировых и белковых (мышечных) запасов. При этом распад структурных белков и депонированного жира сопровождается не только высвобождением необходимой энергии, но и образованием токсичных метаболитов, переводя обменные процессы в стрессовый режим функционирования, а саморегулирующуюся систему организма человека в нестабильное состояние.
 Избыток пищевой энергии – один из основных алиментарных дисбалансов, связанных с употреблением европейского и североамериканского рационов питания. Он обусловлен чрезмерным употреблением всех энергетически ценных макронутриентов (особенно жиров и простых углеводов) и сниженными энергозатратами. Это является причиной развития большого числа алиментарно-зависимых заболеваний.
Избыток пищевой энергии – один из основных алиментарных дисбалансов, связанных с употреблением европейского и североамериканского рационов питания. Он обусловлен чрезмерным употреблением всех энергетически ценных макронутриентов (особенно жиров и простых углеводов) и сниженными энергозатратами. Это является причиной развития большого числа алиментарно-зависимых заболеваний.
Доказано, в частности, что рост числа заболеваний, связанных с избыточной массой тела и ожирением, атеросклерозом, гипертонической болезнью, снижением толерантности к глюкозе, нарушениями обменных процессов, вторичными иммунодефицитами, обусловлен несколькими внешними факторами, ведущими из которых является быстрое неэволюционное изменение питания (соотношение основных групп нутриентов) и образа жизни (снижение физической активности и усиление стресса). Человек в течение многотысячелетней эволюции адаптировался к поступлению определенной пищи в составе рациона. Химический состав рациона и соотношения в нем основных групп нутриентов во многом определили метаболический профиль современного человека. Традиционный среднеевропейский рацион, базирующийся на растительных продуктах с низким содержанием сахара и жира, за последние 50-100 лет практически мгновенно (по эволюционным меркам) существенно изменился. Он стал содержать примерно в два раза больше жира (главным образом за счет животного), в 10-20 раз больше моно- и дисахаридов при снижении в 2-3 раза количества крахмальных и некрахмальных полисахаридов, кальция, многих витаминов. Сложившаяся ситуация имеет устойчивую тенденцию к развитию как по увеличению калорийности, так и по снижению доли растительных источников продовольствия. Пищевой рацион должен содержать 80-90 г белков, 100-150 г жиров, 360-400 г углеводов, энергетическая ценность его должна составлять 2750-2800 ккал.
Второй закон термодинамики в открытых системах
Закрытые термодинамические системы стремятся перейти в состояние термодинамического равновесия, которое характеризуется прекращением в системе всех макроскопических процессов. В таком состоянии система может оставаться сколь угодно долго без внешних воздействий. Открытые системы также стремятся перейти в состояние равновесия, однако постоянный обмен Е и веществом со средой не позволяет этого. Если этот обмен сбалансирован, биосистема находится в состоянии динамического равновесия, т.е. экзергонические и эндоргонические реакции, анаболизм и катаболизм, ассимиляция и диссимиляция уравновешивают друг друга.
2 закон термодинамики – закон вероятности[5] и направленности термодинамического процесса: самопроизвольно теплота переходит лишь от нагретого тела к более холодному, от вещества с большей концентрацией в сторону меньшей. Иными словами, невозможны такие процессы, единственным результатом которых являлось бы отъятие от некоторого тела определенного количества теплоты и превращение этого тепла полностью в работу. Часть тепла всегда будет передаваться более холодному телу. Второй закон показывает ограниченность превращения энергии в работу (КПД<100%) и необратимость термодинамических процессов в открытых системах[6].
Иными словами, все вещества, все виды энергии стремятся перейти в устойчивое состояние, т.е. распределиться в пространстве максимально равномерно. Движущую силу, направленную в область, где значение параметра меньше, называют градиентом (например, концентрационный градиент). Градиент равен отношению разности величин параметра в двух точках к расстоянию между ними[7]. Существование градиента свободной энергии говорит о неравновесности системы, энергия будет перемещаться из области большей в область меньшей, движение энергии будет упорядочено, а энтропия низкая. Вероятность существования такой системы очень мала.
Закрытые системы подчиняются закону увеличения энтропии. При самопроизвольном превращении энергии системы теряют разность энергетических уровней и утрачивают способность совершать работу (по мере использования градиенты уменьшаютсяJ, разница между "больше" и "меньше" исчезает).Поэтому тепловая энергия системы при самопроизвольных процессах возрастает и достигает максимума, когда система приходит в истинное равновесие (тепловая смертьJ). Энтропия (от греч. превращение) – мера утраты способности системы совершать работу, мера рассеяния энергии: S = Q/T, Q = ST. Иными словами, полезная энергия + энтропия = constanta[8].
Без вмешательства извне E→ 0, Э → ¥. Чтобы "всеобщего равенства" не наступило, клетки создают и поддерживают градиенты (например, работа Na+/K+- насоса), затрачивая на это энергию, полученную из внешней среды.
Совершая непрерывную работу, живые системы постоянно расходуют энергию, но при этом живой организм удерживает «неравновесность» – разницу энергетических уровней[9], что возможно благодаря поступлению в нашу систему веществ из окружающей среды, которые имеют большой запас свободной энергии. Поэтому понятие энтропии для живых организмов было заменено понятием потока энтропии. Поступление питательных веществ в организм рассматривается как поток отрицательной энтропии (негэнтропии), поскольку при ассимиляции происходит повышение упорядоченности живой системы. Энергия, которая уходит в ходе совершения работы, представляет собой поток положительной энтропии.
Может показаться, что живые существа не подчиняются второму закону термодинамики, потому что в них S не увеличивается, а находится на постоянном уровне. Но следует учитывать, что они – открытые системы. С этой точки зрения уменьшение S и увеличение E фотосинтезирующих организмов происходит благодаря уменьшению E и увеличению S в системе Солнце-Земля. Точно так же уменьшение E в частях клетки, где идет биохимический синтез, происходит за счет избыточного увеличения S в реакциях диссимиляции, и общий баланс S увеличивается. Поэтому живые организмы подчиняются второму началу термодинамики, когда речь идет о круговороте и превращениях свободной энергии в системе организм – окружающая среда.
Стационарное состояние
Поток отрицательной энтропии возникает в процессе ассимиляции, а положительной – в процесс диссимиляции, что позволяет нам поддерживать динамическое равновесие. Процессы обмена протекают в 2-х направлениях: 1. Самопроизвольно, без дополнительных затрат Е, происходят лишь пассивные процессы, связанные с увеличением Э. 2. Активные процессы, требующие затрат Е. Если эти два потока уравновешивают друг друга, достигается стационарное состояние[10]. Живой организм в каждый момент времени не отвечает приведенному определению стационарного состояния. Однако, если рассмотреть средние значения его параметров за сравнительно большой промежуток времени, можно отметить их постоянство (температура определенных органов и тканей у теплокровных, солевой состав, осмотическое давление и водородный показатель (рН) различных биологических жидкостей. Поддерживаемые на одном уровне параметры называют константами гомеостаза.
| Термодинамическое равновесие | Стационарное состояние |
| Отсутствие обмена с окружающей средой веществом и энергией | Непрерывный обмен с окружающей средой веществом и энергией |
| Энтропия системы постоянна и соответствует максимально возможному в данных условиях значению | Энтропия системы постоянна, но не равна максимально возможному в данных условиях значению. |
| Полное отсутствие в системе каких-либо градиентов | Наличие постоянных по величине градиентов |
| Не требуется затраты свободной энергии | Необходимы постоянные затраты энергии |
| Система нереакционноспособна и не совершает работу против внешних сил | В системе совершаются необратимые реакции, ее работоспособность постоянна и не равна нулю |
Уровни стационарных состояний в течение онтогенеза постоянно изменяются. Живой организм может изменить уровень стационарного состояния в результате воздействия окружающей среды и при патологических процессах.
Одной из важнейших характеристик биологических систем является устойчивость стационарных состояний. Устойчивое стационарное состояние характеризуется тем, что при отклонении системы от стационарного уровня в ней возникают силы, стремящиеся вернуть ее в первоначальное положение. Внешние воздействия вызывают в неустойчивой стационарной системе нарастающие изменения, в результате которых система переходит или в новое устойчивое стационарное состояние (при дополнительной затрате энергии), или в состояние термодинамического равновесия.
 Диаграмма устойчивого (а) и неустойчивого (b) стационарных состояний в открытой системе.
Диаграмма устойчивого (а) и неустойчивого (b) стационарных состояний в открытой системе.
Если построить график зависимости Т∙dS/dt от h (где Т - температура, dS/dt - скорость производства энтропии, h - какой-либо показатель стационарного уровня системы), то для устойчивого стационарного состояния график будет представлен в виде параболы, ветви которой направлены вверх. При неустойчивом стационарном состоянии ветви параболы направлены вниз. Точка А является наиболее устойчивой. Если поместить шарик во внутрь параболы, то его положение наиболее устойчиво. Таким образом, устойчивое стационарное состояние характеризуется тем, что система не может самопроизвольно выйти из него за счет внутренних изменений (шарик из точки А не может скатиться в точку В самопроизвольно, для этого нужно совершить работу). Любое отклонение от уровня стационарности вызовет в системе увеличение скорости продуцирования энтропии. Для точки А скорость производства энтропии будет меньше, чем для точки В. Точка Ai - наиболее неустойчивое состояние системы, так как под влиянием любого внешнего толчка шарик, помещенный в эту точку, быстро удаляется от нее.
И. Пригожин (1946) на основе изучения открытых систем сформулировал основное свойство стационарного состояния: в стационарном состоянии при фиксированных внешних параметрах скорость продукции энтропии в открытой системе, обусловленная протеканием необратимых процессов, постоянна во времени и минимальна по величине. Организм стремится работать в самом выгодном энергетическом режиме. При этом энтропия возрастает с минимальной скоростью. Например, температура тела человека 310 К, а среды – 303 К, и возможность совершения работы за счет тепловой энергии 2 %. Но человек используее для совершения работы химическую энергию, что позволяется увеличить КПД. КПД биологических процессов: гликолиз – 36%, окислительное фосфорилирование – 55%, фотосинтез – 75%, сокращение мышц – 40%, свечение бактерий – до 96%. Наш организм стремится к созданию постоянной величины энтропии, но эмоциональная нагрузка и заболевания приводят к нарушению потоков энтропии, что выводит нас из состояния равновесия.
Стационарное состояние живого организма характеризуется высокой динамичностью: в организме протекают такие процессы, которые нарушают стационарное состояние тех или иных систем (повышение температуры тела при воспалительных процессах). Если эти изменения не превышают определенных пределов, то стационарное состояние может восстановиться, т. е. небольшие отклонения от стационарного состояния не ведут к необратимым изменениям. Биологические системы обладают способностью к саморегуляции (установление наиболее экономичных, наиболее надежных уровней работы). Это положение впервые было сформулировано в работе Ле-Шателье.
Принцип автостабилизации биологических систем: Всякая система, находящаяся в состоянии равновесия и отклонившаяся от этого состояния под воздействием внешнего возмущения, стремится самопроизвольно вернуться в равновесное состояние за счет изменения параметров в направлении, противоположном тому, которое вызвало возмущение. Стремление энтропии в стационарном состоянии к минимальной величине приводит к тому, что при отклонениях от стационарного уровня в системе наступают такие изменения, которые стремятся вернуть ее к минимуму производства энтропии. Принцип Ле-Шателье является следствием закона сохранения энергии, лежит в основе регулирования по типу обратных связей и поддержания гомеостаза в организме.
Мера сложности и самоорганизации связана с понятием информации. Иными словами, информация – это коммуникация и связь, в процессе которой устраняется неопределенность (У. Эшби), или это обозначение содержания, полученного от внешнего мира, и процесса приспособления к нему (Н. Виннер). Информация = мера неоднородности распределения материи и энергии в пространстве и времени, мера изменений, которми сопровождаются все протекающие в мире процесс (В.М. Глушков). Информация – это всеобщее свойство материи, заключающееся в способности материальных объектов сохранять следы былых взаимодействий. Информация проявляется через отражение и может быть оценена количественно [из Игнатовой]. Т.е. биоразнообразие, генетическая, иммунологическая, нейрологическая память, речь, письменность, интернет – это разновидность накопления и передачи информации в живх системах, способствующие самоорганизации, саморегуляции, поддержанию негэнтропии.
3 начало термодинамики. Энтропия любой систем стремится к конечному пределу, не зависящему от давления, плотности или фаз, при стремлении температур к абсолютному нулю J.
Дата добавления: 2018-02-18; просмотров: 2675; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!