ЗАДАЧИ ДЛЯ САМОСТОЯТЕЛЬНОГО РЕШЕНИЯ
Тема 7. Теория гена
Ген – это структурная единица наследственной информации далее неделимая в функциональном отношении. Впервые термин «ген» ввел В.Л. Иогансен в 1909 году.
Существование гена, как дискретной единицы наследственной информации было постулировано Г. Менделем. Первая успешная попытка формирования представлений о гене была осуществлена в работах школы Т. Морганом с дрозофилой. Представления Т. Моргана о гене были изложены в классическом труде «Теория гена», вышедшего в 1926 г. Основные положения этой теории сводятся к следующему:
- Все признаки организмов находятся под контролем генов;
- Гены находятся в хромосомах;
- Гены представляют собой далее не делимые единицы мутации, рекомбинации и функции.
Таким образом, по мнению Т. Моргана ген является единицей:
1) мутации;
2) рекомбинации;
3) функции.
Для определения аллельности двух независимо возникших мутации, т.е. затрагивают один и тот же или разные гены, Т. Морган разработал критерии аллелизма. Он предложил два критерия:
- рекомбинационный – если мутации не рекомбинируют, то они аллельны;
- функциональный – если при скрещивании двух мутантов появляется гибрид дикого типа, то мутации затрагивают разные функции, если образуется мутант, то мутации затрагивают одну и ту же функцию (рис.1)
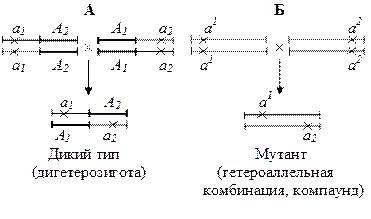
Рис.1. Функциональный критерий аллелизма. А – мутации в разных генах; Б – мутации в одном гене. Крестиками помечены мутации.
|
|
|
Дальнейший вклад в развитие теории гена внесли А.С. Серебровский и его сотрудники. В 1929-1930 гг. они впервые экспериментально доказали сложную структуру гена в работах по исследованию ступенчатого аллеломорфизма. Это явление было открыто при исследовании локуса sc-ac, который контролирует развитие щетинок на теле дрозофилы. Мутации в этом локусе приводят к редукции разных щетинок. Под действием Х-лучей А.С. Серебровским и его сотрудниками был получен ряд мутантов по этому локусу. При скрещивании таких мутантов между собой у гетерозигот, как правило, отсутствовали лишь те щетинки, которые были редуцированы у обеих гомозиготных родительских особей. Так, если в результате мутации sc 1 редуцируются щетинки в участке ABC, а в результате мутации sc 2 редуцируются щетинки в участке BCD, то при скрещивании таких особей между собой у потомства в компаунде будут отсутствовать щетинки только в участке BC. При графическом изображении результатов скрещивания нескольких пар аллеломорфов получалась лестница, ступенями которой служили отдельные аллели (рис.2). Это явление получило название «ступенчатый аллеломорфизм»
| sc 1 | 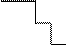 ABC ABC
|
| sc2 | BCD |
| sc3 | CDE |
Рис.2. Графическое изображение результатов скрещивания
|
|
|
нескольких пар аллеломорфов.
Н.П. Дубининым был сделан вывод о том, что ген sc состоит из более мелких элементов – центров и при мутировании изменяется не весь ген, а только его центры.
Изучение ступенчатого аллеломорфизма показало, что ген не является единицей мутации и аллельные мутации способны рекомбинировать.
В 40-х годах прошлого века начали появляться работы М. Грина, К. Оливера, Е. Льюиса в которых была показана рекомбинация мутаций, которые в соответствии с функциональным тестом Моргана должны быть аллельными. Это противоречие между рекомбинационным и функциональным критерием аллелизма и отразилось в термине «псевдоаллелизм». Вначале предполагали, что псевдоаллелизм встречается только в исключительных случаях, но затем, с увеличением числа анализируемых особей в выборках, стало очевидным широкое распространение данного явления.
Большой вклад в развитие теории гена внесли Дж. Бидл и Е. Тейтум. В своей работе, вышедшей в 1941 г., они сформулировали концепцию «один ген – один фермент».
Дж. Бидл и Э. Тейтум облучали микроконидии нейроспоры рентгеновскими лучами, а затем высевали их на полноценную питательную среду. Затем выросшие колонии пересевали на минимальную среду. Показано, что некоторые колонии (мутантные) не способны расти на минимальной среде.
|
|
|
Дж. Бидл и Э. Тейтум установили, что каждая мутация блокирует синтез какой-либо вещества на одном из этапов. Эти мутации можно разбить на группы соответственно числу стадий в синтезе данного соединения. При этом мутации каждой группы аллельны друг другу, т. е. при скрещивании между собой они будут образовывать мутантные гибриды, тогда как при скрещивании мутантов из разных групп будут образовываться гибриды дикого типа.
Огромным вкладом в развитие теории гена стали исследования С. Бензером локуса rII бактериофага Т4 Е. coli . Мутанты бактериофага Т4 по локусу rII образуют на газоне Е. coli В более крупные стерильные пятна, по сравнению с фагом дикого типа. Отличительной особенностью этих мутантов, по сравнению с диким типом, является также то, что они не могут размножаться в клетках штамма Е. coli К12, лизогенного по фагу λ (табл2).
Таблица 2
Тип колоний, образуемых фагом Т4 и мутантом Т4
по локусу rII на газоне Е. coli
| Штамм | Генотип Т4 | |
| Е. coli | Дикий тип | Мутант rII |
| В |  мелкие
мелкие
| крупные |
| К12 | мелкие | Не образует |
Если фаголизатом, полученным при заражении клеток Е. coli В двумя разными мутантами бактериофага Т4 по локусу rII , инфицировать клетки Е. coli К-12 на газоне появляются бляшки дикого типа. Таким образом, в ходе репликации фаговых геномов после инфекции бактериального штамма двумя мутантами rII могут образовываться рекомбинанты дикого типа. Частота появления этих рекомбинантов пропорциональна расстоянию между мутантными точками в геноме бактериофага.
|
|
|
Сложные аллельные отношения, возникающие при скрещивании двух мутантов и приводящие к появлению рекомбинантов в 1957 г. получили название межаллельная комплементация. Матрица межаллельной комплементации включает результаты всех проведенных скрещиваний. Если при скрещивании мутантов образуется гибрид дикого типа, то в соответствующей строке ставят знак «+», если образуется мутант – ставят знак «-».
На основе функционального теста на аллелизм выяснилось, что локус rII включает два гена: А и В. В пределах этих генов было картировано более 2000 мутантов. Бензер показал, что рекомбинация происходила на расстоянии около двух нуклеотидных пар и, очевидно, что рекомбинация может разделять соседние пары нуклеотидов, что наименьший участок ДНК, который изменяется при мутировании, – это пара нуклеотидов.
Для точной локализации мутации Бензер использовал не только скрещивание точковых мутантов между собой, но и метод перекрывающихся делеций. При скрещивании точкового и делеционного мутантов рекомбинанты появляются только тогда, когда делеция не затрагивает участок, в котором локализована точковая мутация.
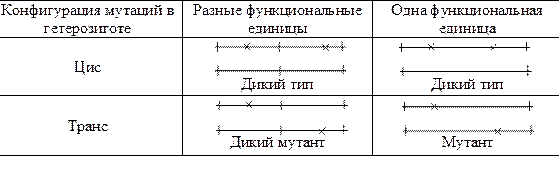 С. Бензер попытался ревизовать и понятие «ген». Для выяснения затрагивают ли две мутации одну и туже или разные единицы функции он предложил использовать цис-транс-тест, изобретенный Е. Льюисом. Согласно этому тесту мутации попарно испытывают в гетерозиготе в двух конфигурациях: цис – когда обе мутации в гибриде происходят от одного родителя, и транс – когда они поступают в гибрид от разных родителей (рис.3). Согласно С. Бензеру, если цис- и транс-гетерозигота имеют одинаковый (дикий) фенотип, то мутации затрагивают разные единицы функций, а если цис- и транс-гетерозиготы разного фенотипа (цис – дикий, а транс – мутантный), то мутации затрагивают одну единицу функции, которую он предложил называть цистроном.
С. Бензер попытался ревизовать и понятие «ген». Для выяснения затрагивают ли две мутации одну и туже или разные единицы функции он предложил использовать цис-транс-тест, изобретенный Е. Льюисом. Согласно этому тесту мутации попарно испытывают в гетерозиготе в двух конфигурациях: цис – когда обе мутации в гибриде происходят от одного родителя, и транс – когда они поступают в гибрид от разных родителей (рис.3). Согласно С. Бензеру, если цис- и транс-гетерозигота имеют одинаковый (дикий) фенотип, то мутации затрагивают разные единицы функций, а если цис- и транс-гетерозиготы разного фенотипа (цис – дикий, а транс – мутантный), то мутации затрагивают одну единицу функции, которую он предложил называть цистроном.
Рис.3. Схема цис-транс-теста. Крестики – мутации.
Если ген кодирует синтез фермента какого-либо биосинтетического пути (см. схему на рис. 4), то мутация в гене (например, гене с) может вызвать инактивацию фермента, и, соответственно, блокировать синтез остальных промежуточных продуктов данного пути (соединений IV–VI).
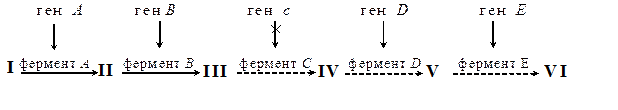 |
Рис. 4. Схема биосинтетического пути
РЕШЕНИЕ ТИПОВЫХ ЗАДАЧ
Задача 1. У бактерий Escherichia coli было получено 5 мутантов, затрагивающих разные этапы одного и того же биосинтетического пути (схема пути приведена на рис. 4), и изучена их способность к росту на среде, содержащей соединения (I–V), являющиеся предшественниками конечного продукта VI (табл. 3).Вам необходимо определить правильную очередность синтеза соединений I-VI в рассматриваемом биосинтетическом пути, а также идентифицировать мутанты, т.е. определить этап, где произошла мутация у каждого из них.
Таблица 3
Характеристика мутантов Escherichia coli
| Мутанты | Соединения | |||||
| I | II | III | IV | V | VI | |
| 1 | - | - | - | + | - | + |
| 2 | - | + | - | + | - | + |
| 3 | - | - | - | - | - | + |
| 4 | - | + | + | + | - | + |
| 5 | + | + | + | + | - | + |
«+» – есть рост, «-» – нет роста.
Решение:
При решении задач подобного типа следует помнить, что если исследуемый ауксотрофный мутант растет на среде только в случае добавки необходимого метаболита, то у него блокирован какой-то этап биосинтетического пути на этапе, предшествующем этому метаболиту, то есть метаболит занимает в цепи место после мутационного блока. Другие более ранние предшественники, находящиеся в биосинтетическом пути перед блоком, не могут обеспечить рост мутанта. С другой стороны, если мутант накапливает определенный метаболит, то у него блокирован этап пути биосинтеза сразу же после накапливаемого метаболита. Например, в случае соединения V мы видим, что оно не обеспечивает рост ни одного из мутантов. Следовательно, в рассматриваемом биосинтетическом пути соединение V является самым первым предшественником. Затем следует соединение I, которое обеспечивает рост только одного мутанта (а именно, мутанта № 5), значит, в биосинтетическом пути оно располагается следующим. Затем следует соединение III и т.д. Следовательно, очередность синтеза соединений I-VI должна быть следующей: V ® I ® III ® II ® IV ® VI.
Подобным образом рассуждаем и в случае идентификации мутантов. Поскольку мутант 5 не растет на соединении V, значит, мутация у него произошла на участке V ®I. Мутант 4 не растет уже на двух соединениях – V и I, следовательно, мутация у него произошла на следующем участке между I ®III. Мутант 2 не может расти на соединениях V, I и III, следовательно, мутация у него произошла на участке III ®II. Рассуждая подобным образом и далее, определяем местоположение всех остальных мутаций.
Ответ.
Мутанты: 5 4 2 1 3
 Путь: V I III II IV VI.
Путь: V I III II IV VI.
Такой принцип идентификации мутантов применим для любого биосинтетического пути, и является универсальным как для прокариотических, так и эукариотических организмов.
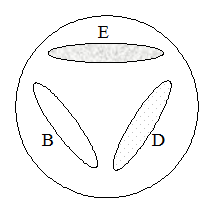 Задача 2. Три независимых триптофан-зависимых мутанта дрожжей, обозначенных trp B, trp D и trp E, соответственно, были ресуспендированы в физиологическом растворе и нанесены на агаризованную среду без триптофана в виде полосок, таким образом, что отдельные штрихи не соприкасались между собой, но могли использовать для своего роста метаболиты других мутантов (рис. 5). После выращивания мутантов в течение 2-3 суток был зарегистрирован рост дрожжевых клеток, как указано на рисунке. Проанализируйте результаты эксперимента и определите, какова очередность действия генов триптофанового пути у дрожжей?
Задача 2. Три независимых триптофан-зависимых мутанта дрожжей, обозначенных trp B, trp D и trp E, соответственно, были ресуспендированы в физиологическом растворе и нанесены на агаризованную среду без триптофана в виде полосок, таким образом, что отдельные штрихи не соприкасались между собой, но могли использовать для своего роста метаболиты других мутантов (рис. 5). После выращивания мутантов в течение 2-3 суток был зарегистрирован рост дрожжевых клеток, как указано на рисунке. Проанализируйте результаты эксперимента и определите, какова очередность действия генов триптофанового пути у дрожжей?
|
Учитывая, что наличие роста на конкретном соединении говорит о том, что блок биосинтетического пути находится на более раннем его этапе, производим ана-
лиз картины роста мутантов дрожжей. Мы видим, что мутант trp E растет хорошо, и, следовательно, способен использовать для роста метаболиты мутантов trp D и trp B.
Значит в пути синтеза триптофана, фермент, кодируемый геном trp E, действует первым. Рост мутанта trp D слабый и неравномерный, этот говорит о том, что только один из двух мутантов обеспечивает его необходимым фактором роста. Поскольку мутант trp E имеет самый «ранний» блок триптофанового пути и не способен синтезировать метаболиты для «подпитки» других мутантов, следовательно, только клетки мутанта trp B синтезируют нужное для его роста соединение. Действительно, pост клеток мутантa trp D является более обильным в участке, приближенном к месту посева мутанта trp B. Значить фермент, кодируемый геном trp D, работает следующим.
Ответ. Последовательность генов в биосинтетическом пути:
trp E→ trp D → trp B.
Задача 3. В таблице приведены результаты теста на комплементарность для десяти точковых мутаций. «+» – комплементация мутации; «-» – отсутствие комплементации. По результатам, приведенным в таблице, определите группы комплементации.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
| 1 | - | + | + | + | + | - | - | + | + | + |
| 2 | - | + | + | + | + | + | + | + | - | |
| 3 | - | - | - | + | + | - | - | + | ||
| 4 | - | - | + | + | - | - | + | |||
| 5 | - | + | + | - | - | + | ||||
| 6 | - | - | + | + | + | |||||
| 7 | - | + | + | + | ||||||
| 8 | - | - | + | |||||||
| 9 | - | + | ||||||||
| 10 | - |
Решение:
1). Анализируем первую горизонтальную строку. Мы видим, что при скрещивании первого мутанта с шестым и седьмым мутантом не происходит комплементации мутации (знак «-»), следовательно, эти мутации затрагивают одну область гена, поэтому мы можем отнести эти мутации к одной группе комплементации. При скрещивании первого мутанта со вторым, третьим, четвертым, пятым, восьмым, девятым и десятым мутантом происходит комплементации мутации (знак «+»), следовательно, эти мутации затрагивают разные области гена, поэтому мы можем отнести эти мутации к другой группе комплементации.
2). Анализируем вторую горизонтальную строку. Мы видим, что при скрещивании второго мутанта с мутантом десять не происходит комплементации мутации (знак «-»), следовательно, эти мутации затрагивают одну область гена, поэтому мы можем отнести эти мутации к одной группе комплементации.
3). Анализируем третью горизонтальную строку. Мы видим, что при скрещивании третьего мутанта с мутантами четыре, пять, восемь и девять не происходит комплементации мутации (знак «-»), следовательно, эти мутации затрагивают одну область гена, поэтому мы можем отнести эти мутации к одной группе комплементации.
Ответ: Группа 1: мутанты 1, 6, 7.
Группа 2: мутанты 2, 10.
Группа 3: мутанты 3, 4, 5, 8, 9.
Задача 4. В результате эксперимента, получено пять делеционных варианта гена Х и пять точковых мутанта этого гена. Карта делеций представлена на рис.6. Пять точковых мутанта скрестили с делеционными вариантами. Результаты скрещивания представлены в таблице.
| 1 | |||||

| 2 | ||||
| 3 | |||||
| 4 | |||||
| 5 | |||||
Рис.6. Делеционные варианты гена Х
| Делеция | Точковый мутант | ||||
| а | b | c | d | e | |
| 1 | - | - | - | - | - |
| 2 | - | - | + | - | - |
| 3 | - | - | + | + | - |
| 4 | + | - | + | + | - |
| 5 | + | - | + | + | + |
Решение:
Как указывалось выше, при скрещивании точкового и делеционного мутанта рекомбинанты появляются только тогда, когда делеция не затрагивает участок, в котором локализована точковая мутация. Делеции разделяют карту гена Х на пять областей (рис.6). Если при скрещивании точкового мутанта происходит рекомбинация только с некоторыми делециями, то эта точковая мутация локализована слева от самой протяженной делеции с которой произошла рекомбинация.
1. Анализируем результаты скрещивания мутанта а. Как видно из первого столбца таблицы при скрещивании точкового мутанта а с делеционными мутантами 4 и 5 происходит рекомбинация (знак «+»), а при скрещивании с делециями 1, 2 и 3.рекомбинация не происходит (знак «-»). Следовательно, данная точковая делеция локализована в области 3.
2. Анализируем результаты скрещивания мутанта b. Как видно из второго столбца таблицы при скрещивании точкового мутанта b с делеционными мутантами не происходит рекомбинация не с одной делецией (знак «-»). Следовательно, данная точковая делеция локализована в области 5.
3. Анализируем результаты скрещивания мутанта с. Как видно из третьего столбца таблицы при скрещивании точкового мутанта с с делеционными мутантами 2, 3, 4 и 5 происходит рекомбинация (знак «+»), а при скрещивании с делецией 1 рекомбинация не происходит (знак «-»).Следовательно, данная точковая делеция локализована в области 1.
4. Анализируем результаты скрещивания мутанта d. Как видно из таблицы при скрещивании точкового мутанта d с делеционными мутантами происходит рекомбинация мутации с делециями 3, 4 и 5 (знак «+»), но не рекомбинирует с делециями 1 и 2 (знак «-»). Следовательно, данная точковая делеция локализована в области 2.
5. Анализируем результаты скрещивания мутанта е. Как видно из таблицы мутант е рекомбинирует с делецией 5, но не рекомбинирует с делециями 1, 2, 3 и 4. Следовательно, данная точковая делеция локализована в области 4.
Ответ:
| 1 | 2 | 3 | 4 | 5 | |
| c | d | а | e | b |
Задача 5. В таблице представлены результаты попарных скрещиваний между пятью rII-делециями. «+» – комплементация мутации; «-» – отсутст-вие комплементации. Постройте карту делеций с указанием относительной длинны каждой из них. Какие из этих мутаций могли бы быть точковыми?
| 1 | 2 | 3 | 4 | 5 | |
| 1 | - | - | - | - | + |
| 2 | - | - | + | + | |
| 3 | - | + | + | ||
| 4 | - | - | |||
| 5 | - |
Решение:
1). Анализируем графы таблицы для определения самой протяженной делеции. Как видно из таблицы при скрещивании первого делеционного варианта с другими делециями комплементация происходит только при скрещивании с делецией 5 (знак «+»), следовательно, делеция 1 является самой протяженной.
2). Наносим на карту делецию которая комплементирует мутацию при скрещивании с самой протяженной. Это делеция 5. Отмечаем делецию 5 справа или слева от делеции 1.
3). При скрещивании делеции 2 с делецией 1 не происходит комплементации мутации (знак «-»), следовательно, она затрагивает делецию 1, но при скрещивании делеции 2 с делецией 5 происходит комплементация мутации (знак «+»), следовательно, она не затрагивает делецию делецию 5.
4). При скрещивании делеции 3 с делециями 1 и 2 не происходит комплементации мутации (знак «-»), следовательно, она затрагивает делецию 1 и 2, но при скрещивании делеции 3 с делецией 5 происходит комплементация мутации (знак «+»), следовательно, она не затрагивает делецию делецию 5.
5). При скрещивании делеции 4 с делецией 1 и 5 не происходит комплементации мутации (знак «-»), следовательно, она затрагивает эти делеции, но при скрещивании делеции 4 с делециями 2 и 3 происходит комплементация мутации (знак «+»), следовательно, она не затрагивает делеции 2 и 3.
Ответ. Карта rII-делециями делеций:
 1
1
2
3 5
4
Точковыми могли бы быть мутации 5, 3, 2.
Задача 6. У Pseudomonas putida получены ауксотрофные мутанты дефектные по различным этапам синтеза триптофана. Используя характеристику мутантов, приведенную в таблице, определите последовательность стадий биосинтеза триптофана у этих бактерий.
| Рост на средах с предшественником | |||||
| хоризмат | антранилат | КДРФ | индол | триптофан | |
| trpB | - | - | - | - | + |
| trpE | - | + | + | + | + |
| trpC | - | - | - | + | + |
| trpA | - | - | - | + | + |
| trpD | - | - | + | + | + |
Решение:
Если мутант не растет на минимальной среде, а способен к росту только при внесении в среду определенного предшественника синтеза триптофана, то у него блокирован этап метаболической цепи перед биосинтезом этого метаболита.
1.Мутант trpE растет при добавлении в среду любого предшественника, кроме хоризмата, следовательно, у него блокирован самый ранний этап синтеза триптофана сразу после синтеза хоризмата.
2.Мутант trpD способен расти на средах со всеми промежуточными продуктами синтеза триптофана, кроме антранилата и КДРФ. Это означает, что мутационный блок располагается сразу после синтеза антранилата, который в цепи биосинтеза находится сразу после хоризмата.
3.Мутант trpC способен использовать для своего роста триптофан, и индол, но не антраниловую кислоту и КДРФ. Следовательно, мутация trpC прерывает цепь биосинтеза сразу после КДРФ, который располагается в цепи синтеза после антраниловой кислоты, но перед индолом и триптофаном.
4. Мутант trpA растет только на среде с триптофаном и индолом. Можно заключить, что цепь биосинтеза у него блокирована на этапе между индол-3-глицерофосфатом и индолом.
5. Мутант trp В растет только на среде с триптофаном. Можно заключить, что цепь биосинтеза у него блокирована на этапе между индолом и триптофаном.
Таким образом, последовательность метаболитов в цепи биосинтеза триптофана следующая: хоризмовая кислота → антраниловая кислота → КДРФ →индол → индол-3-глицерофосфат → триптофан.
ЗАДАЧИ ДЛЯ САМОСТОЯТЕЛЬНОГО РЕШЕНИЯ
1. В таблице приведены результаты теста на комплементарность для десяти точковых мутаций. «+» – комплементация мутации; «-» – отсутствие комплементации. По результатам, приведенным в табл., определите группы комплементации.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| 1 | - | + | + | - | + | + | + |
| 2 | - | - | + | - | + | + | |
| 3 | - | - | - | + | + | ||
| 4 | - | + | + | + | |||
| 5 | - | + | + | ||||
| 6 | - | - | |||||
| 7 | - |
2. В таблице приведены результаты теста на комплементарность для десяти точковых мутаций. «+» – комплементация мутации; «-» – отсутствие комплементации. По результатам, приведенным в таблице, определите группы комплементации.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
| 1 | - | + | + | + | + | + | + | - | - |
| 2 | - | + | + | + | + | - | + | + | |
| 3 | - | - | - | - | + | + | + | ||
| 4 | - | - | - | + | + | + | |||
| 5 | - | - | + | + | + | ||||
| 6 | - | + | + | + | |||||
| 7 | - | + | + | ||||||
| 8 | - | - | |||||||
| 9 | - |
3. В таблице приведены результаты теста на комплементарность для десяти точковых мутаций. «+» – комплементация мутации; «-» – отсутствие комплементации. По результатам, приведенным в таблице, определите группы комплементации.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| 1 | - | + | - | + | + | + | - |
| 2 | - | + | - | + | - | + | |
| 3 | - | + | + | + | - | ||
| 4 | - | + | - | + | |||
| 5 | - | + | + | ||||
| 6 | - | + | |||||
| 7 | - |
4. В таблице приведены результаты теста на комплементарность для десяти точковых мутаций. «+» – комплементация мутации; «-» – отсутствие комплементации. По результатам, приведенным в таблице, определите группы комплементации.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| 1 | - | + | + | + | + | + | + | + |
| 2 | - | + | + | - | + | + | - | |
| 3 | - | - | + | - | - | + | ||
| 4 | - | + | - | - | + | |||
| 5 | - | + | + | - | ||||
| 6 | - | - | + | |||||
| 7 | - | + | ||||||
| 8 | - |
5. В таблице приведены результаты теста на комплементарность для десяти точковых мутаций. «+» – комплементация мутации; «-» – отсутствие комплементации. По результатам, приведенным в таблице, определите группы комплементации.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
| 1 | - | + | + | + | - | + | + | + | - | + | + | - |
| 2 | - | + | - | + | - | + | + | + | - | + | + | |
| 3 | - | + | + | + | - | - | + | + | - | + | ||
| 4 | - | + | - | + | + | + | - | + | + | |||
| 5 | - | + | + | + | - | + | + | - | ||||
| 6 | - | + | + | + | - | + | + | |||||
| 7 | - | - | + | + | - | + | ||||||
| 8 | - | + | + | - | + | |||||||
| 9 | - | + | + | - | ||||||||
| 10 | - | + | + | |||||||||
| 11 | - | + | ||||||||||
| 12 | - |
6. В результате эксперимента, получено пять делеционных варианта гена Х и пять точковых мутанта этого гена. Карта делеций представлена на рис.7. Пять точковых мутанта скрестили с делеционными вариантами. Результаты скрещивания представлены в таблице.
| D1 | |||||
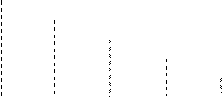
| D2 | ||||
| D3 | |||||
| D4 | |||||
| D5 | |||||
Рис.7. Делеционные варианты гена Х
| Делеция | Точковый мутант | ||||
| 1 | 2 | 3 | 4 | 5 | |
| D1 | - | - | - | - | - |
| D2 | + | - | - | - | - |
| D3 | + | - | + | - | - |
| D4 | + | - | + | + | - |
| D5 | + | + | + | + | - |
7. В результате эксперимента, получено семь делеционных варианта гена и семь точковых мутанта этого гена. Карта делеций представлена на рис.8. Пять точковых мутанта скрестили с делеционными вариантами. Результаты скрещивания представлены в таблице.
| D1 | ||||||

| D2 | |||||
| D3 | ||||||
| D4 | ||||||
| D5 | ||||||
| D6 | ||||||
| D7 | ||||||
Рис.8. Делеционные варианты гена
| Делеции | Точковый мутант | ||||||
| а | b | c | d | e | f | g | |
| 1 | - | - | - | - | - | - | - |
| 2 | - | - | - | - | - | - | + |
| 3 | - | - | + | - | + | - | + |
| 4 | + | - | + | - | + | - | + |
| 5 | + | - | + | - | + | - | + |
| 6 | + | - | + | - | + | + | + |
| 7 | + | - | + | + | + | + | + |
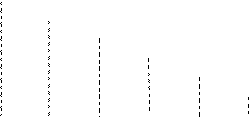
8. В результате эксперимента, получено шесть делеционных варианта гена и семь точковых мутанта этого гена. Карта делеций представлена на рис.9. Семь точковых мутанта скрестили с делеционными вариантами. Результаты скрещивания представлены в таблице.
| D1 | |||||
| D2 | ||||
| D3 | |||||
| D4 | |||||
| D5 | |||||
| D6 | |||||
Рис.9. Делеционные варианты гена
| Делеции | Точковый мутант | ||||||
| а | b | c | d | e | f | g | |
| 1 | - | - | - | - | - | - | - |
| 2 | - | + | - | - | - | + | - |
| 3 | - | + | - | - | + | + | - |
| 4 | - | + | + | - | + | + | - |
| 5 | + | + | + | - | + | + | + |
| 6 | + | + | + | - | + | + | + |
9. В результате эксперимента, получено пять делеционных варианта гена и пять точковых мутанта этого гена. Карта делеций представлена на рис.10. Пять точковых мутанта скрестили с делеционными вариантами. Результаты скрещивания представлены в таблице.
| D1 | ||||

| D2 | |||
| D3 | ||||
| D4 | ||||
| D5 | ||||
Рис.10. Делеционные варианты гена
| Делеция | Точковый мутант | ||||
| 1 | 2 | 3 | 4 | 5 | |
| D1 | - | - | - | - | + |
| D2 | - | + | - | - | + |
| D3 | - | + | - | + | + |
| D4 | + | + | - | + | + |
| D5 | + | + | - | + | + |
10. В результате эксперимента, получено четыре делеционных варианта гена и шесть точковых мутанта этого гена. Карта делеций представлена на рис.11. Шесть точковых мутанта скрестили с делеционными вариантами. Результаты скрещивания представлены в таблице.
| D1 | |||

| D2 | ||
| D3 | |||
| D4 | |||
Рис.11. Делеционные варианты гена
| Делеции | Точковый мутант | |||||
| а | b | c | d | e | f | |
| 1 | + | - | - | - | - | - |
| 2 | + | + | - | - | - | - |
| 3 | + | + | + | - | - | + |
| 4 | + | + | + | + | - | + |
11. В таблице представлены результаты попарных скрещиваний между шестью rII-делециями. «+» – комплементация мутации; «-» – отсутствие комплементации. Постройте карту делеций с указанием относительной длинны каждой из них. Какие из этих мутаций могли бы быть точковыми?
| 1 | 2 | 3 | 4 | 5 | 6 | |
| 1 | - | + | + | - | - | + |
| 2 | - | + | - | - | + | |
| 3 | - | + | + | - | ||
| 4 | - | - | + | |||
| 5 | - | - | ||||
| 6 | - |
12. В таблице представлены результаты попарных скрещиваний между пятью rII-делециями. «+» – комплементация мутации; «-» – отсутствие комплементации. Постройте карту делеций с указанием относительной длинны каждой из них. Какие из этих мутаций могли бы быть точковыми?
| 1 | 2 | 3 | 4 | 5 | |
| 1 | - | + | + | - | - |
| 2 | - | - | - | + | |
| 3 | - | + | + | ||
| 4 | - | - | |||
| 5 | - |
13. В таблице представлены результаты попарных скрещиваний между семью rII-делециями. «+» – комплементация мутации; «-» – отсутствие комплементации. Постройте карту делеций с указанием относительной длинны каждой из них. Какие из этих мутаций могли бы быть точковыми?
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| 1 | - | - | + | - | + | + | + |
| 2 | - | + | + | - | + | + | |
| 3 | - | - | + | - | - | ||
| 4 | - | + | + | + | |||
| 5 | - | + | + | ||||
| 6 | - | + | |||||
| 7 | - |
14. В таблице представлены результаты попарных скрещиваний между шестью rII-делециями. «+» – комплементация мутации; «-» – отсутствие комплементации. Постройте карту делеций с указанием относительной длинны каждой из них. Какие из этих мутаций могли бы быть точковыми?
| 1 | 2 | 3 | 4 | 5 | 6 | |
| 1 | - | + | - | - | + | + |
| 2 | - | - | + | - | + | |
| 3 | - | + | - | + | ||
| 4 | - | + | + | |||
| 5 | - | + | ||||
| 6 | - |
15. В таблице представлены результаты попарных скрещиваний между шестью rII-делециями. «+» – комплементация мутации; «-» – отсутствие комплементации. Постройте карту делеций с указанием относительной длинны каждой из них. Какие из этих мутаций могли бы быть точковыми?
| 1 | 2 | 3 | 4 | 5 | 6 | |
| 1 | - | - | - | + | - | - |
| 2 | - | + | + | + | - | |
| 3 | - | + | + | + | ||
| 4 | - | + | + | |||
| 5 | - | - | ||||
| 6 | - |
16. В норме гаплоидные дрожжи синтезируют пигмент красного цвета. Мутанты имеют различную окраску колоний в зависимости от типа мутации: оранжевую (мутация гена О–), розовую (мутация гена Р–), белую (мутация W–), желтую (мутация Y–) и бежевую (мутация B–). Каждый фенотип является результатом точковой мутации.
Для того чтобы идентифицировать мутанты было проведено скрещивание дрожжей для получения двойных мутантов во всевозможных комбинациях мутаций. Фенотипы двойных мутантов приведены в табл.4.
Таблица 4
Характеристика двойных мутантов дрожжей
| Р¯ | W¯ | Y¯ | В¯ | |
| О¯ Р ¯ W¯ Y ¯ | розовые − | белые белые − | желтые розовые белые − | бежевые розовые белые желтые |
1). Определите последовательность этапов синтеза красного пигмента и расположите гены в порядке контролируемых ими этапов.
2). Определите, в каком соотношении будет образовываться потомство в скрещивании между клетками двойного мутанта О ¯Р¯ и клетками дикого типа О+Р+, если локусы сцеплены и расположены на расстоянии 16 сМ?
17. У штамма N. crassa, клетки которого не способны синтезировать аргинин (Arg-), были получены ревертанты, независимые от аргинина (Arg+). В скрещивании между таким ревертантом и клетками дикого типа было получено потомство (гаплоидное). Какая часть потомства будет аргининнезависимой, если:
1). Ревертанты возникли в результате обратной мутации по тому же самому нуклеотиду (истинная реверсия);
2). Ревертанты возникли в результате мутации в другом гене, локализованном на другой хромосоме;
3). Ревертанты являются результатом мутации в другом гене, который находится на расстоянии 10 сМ от Arg-гена на той же самой хромосоме.
18. Получены различные мутанты бактерий E . coli , нуждающиеся в аспарагиновой кислоте, треонине и метионине. Характеристика мутантов приведена в табл.5.
Таблица 5
Характеристика мутантов E . coli
| Мутант | Необходимый для роста метаболит: | Накапливаемый метаболит | |||||
| аспара-гиновая кислота | гомо-серин | гомо-серин фосфат | треонин | гомо- цисте- ин | метио- нин | ||
| asp A | + | – | – | – | – | – | фумаровая кислота |
| metA | – | – | – | – | + | + | гомосерин |
| metH | – | – | – | – | – | + | гомоцистеин |
| thrC | – | – | – | + | – | – | гомосерин- фосфат |
| thrB | – | – | + | + | – | – | гомосерин |
| thrA | – | + | + | + | – | – | аспарагиновая кислота |
Определите последовательность этапов синтеза соответствующих аминокислот и расположите гены в порядке контролируемых ими этапов.
19. У бактерий E . coli было получено пять мутантов, неспособных синтезировать тимин. Для идентификации мутантов была изучена их способность к росту на средах, содержащих различные предшественники тимина (табл.6).
Таблица 6
Способность к росту мутантов E . coli
на промежуточных продуктах пути синтеза тимина
| Мутанты | Соединения | ||||
| А | В | С | D | Тимин | |
| 9 | + | - | + | - | + |
| 10 | - | - | + | - | + |
| 14 | + | + | + | - | + |
| 18 | + | + | + | + | + |
| 21 | - | - | - | - | + |
1). Определите последовательность этапов синтеза предшест-венников тимина и установите местоположение изучаемых мутаций.
2). Какие предшественники будут накапливаться в клетках мутантов с двойными блоками биосинтетического пути: 9 и 10, 10 и 14?
20. У бактерий Pseudomonas получены ауксотрофные мутанты, дефектные по различным этапам биосинтеза триптофана. Характеристики мутантов приведены в табл.7.
Таблица 7
Характеристика Trp- -мутантов бактерий Pseudomonas
| Мутант | Рост на минимальной среде с добавлением метаболита: | Накапливаемый метаболит | |||
| антранилат | КФАДРФ | индол | триптофан | ||
| trpA | – | – | – | + | индол-3-глице-рофосфат |
| trpB | – | – | – | + | индол |
| trpC | – | – | + | + | КФАДРФ |
| trpD | – | + | + | + | антранилат |
| trpE | + | + | + | + | хоризмат |
Примечание: КФДАРФ – 1-(о-карбоксифениламино)-1-дезоксирибулозо-фосфат.
Определите последовательность этапов синтеза триптофана и расположите гены в порядке контролируемых ими этапов.
21. Предположим, что синтез клеточного метаболита G у бактерий E . coli осуществляется по пути, включающему несколько этапов. Соединения А, B , C , D , E и F являются промежуточными продуктами этого пути, однако порядок их синтеза не известен. Для того чтобы установить очередность синтеза этих соединений, были получены мутанты (номера с 1-го по 7-й) с блоками различных этапов этого пути. Затем мутанты были проверены на способность к росту на среде, содержащей тот или иной предшественник. Ниже в таблице приведены характеристики полученных мутантов (табл.8).
Таблица 8
Способность к росту мутантов E . coli на соединениях А - G
| Мутанты | Соединения | ||||||
| A | B | C | D | E | F | G | |
| 1 | + | + | + | + | + | - | + |
| 2 | - | - | - | - | - | - | + |
| 3 | - | + | + | - | + | - | + |
| 4 | - | + | - | - | + | - | + |
| 5 | + | + | + | - | + | - | + |
| 6 | + | + | + | + | + | + | + |
| 7 | - | - | - | - | + | - | + |
Определите последовательность этапов синтеза промежуточных продуктов А– G в изучаемом биосинтетическом пути и установите местоположение мутаций 1–7.
22. Путь биосинтеза глютамина (Gln) и пролина (Pro) имеет несколько общих промежуточных продуктов. У дрожжей были получены ауксотрофные мутанты 1–7, нуждающиеся для своего роста либо в глутамине, либо пролине. У полученных мутантов также была проверена способность к росту на предшественниках А–Е. Характеристика мутантов представлена в табл.9.
Таблица 9
Способность к росту мутантных дрожжей
на предшественниках синтеза глутамина и пролина
| Мутанты | Соединения | |||||||
| А | В | С | D | E | Gln | Pro | Gln+Pro | |
| 1 | + | - | - | - | + | - | + | + |
| 2 | - | - | - | - | - | - | + | + |
| 3 | - | - | + | - | - | - | - | + |
| 4 | - | - | - | - | - | + | - | + |
| 5 | - | - | + | + | - | - | - | + |
| 6 | + | - | - | - | - | - | + | + |
| 7 | - | + | - | - | - | + | - | + |
Определите последовательность этапов синтеза предшественников глютамина и пролина, а также установите местоположение изучаемых мутаций.
23. Были изолированы мутанты Aspergillus, нуждающиеся для роста на минимальной среде в добавке соединения G. Соединения А-Е являются промежуточными продуктами пути синтеза G , однако порядок их синтеза не известен. Для того, чтобы определить этапы синтеза данного соединения мутанты были проверены на способность к росту на средах в присутствии каждого из этих соединений (табл. 10).
Таблица 10
Способность к росту мутантов дрожжей
на промежуточных продуктах пути синтеза соединения G
| Мутанты | Соединения | |||||
| А | В | С | D | Е | G | |
| 1 | - | - | - | + | - | + |
| 2 | - | + | - | + | - | + |
| 3 | - | - | - | - | - | + |
| 4 | - | + | + | + | - | + |
| 5 | + | + | + | - | - | + |
1) Какова очередность синтеза соединений А- G в рассматриваемом биосинтетическом пути?
2) Определите местоположение мутаций 1-5.
На каких добавках будут расти гетерокарионы, сформировавшиеся у двойных мутантов после скрещивания – 1, 3 и 2, 4; 1, 3 и 3, 4; 1, 2 и 2, 4 и 1, 4?
Дата добавления: 2021-05-18; просмотров: 913; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!