Современные взгляды на развитие человека.
Лекция 62.
Тема: «Этапы ЭВОЛЮЦИИ ЧЕЛОВЕКА»
План.
Этапы эволюции
Методы изучения этапов эволюции
Характеристика основных этапов антропогенеза
1. Обычно выделяют следующие этапы эволюции человека:
1. Древнейшие стадии гоминизации—происхождение рода Homo.
2. Эволюция рода Homo до возникновения современного человека.
3. Эволюция современного человека.
Первый этап антропогенеза есть чисто биологическая эволюция.
На втором этапе к элементарным факторам биологической эволюции подключается действие социального фактора, который на третьем этапе является доминирующим. Методологические подходы к изучению разных этапов антропогенеза различны.
2. Методы изучения этапов эволюции.
Изучение 1-го этапа производят методами палеонтологии и сравнительной анатомии. В связи с появлением элементов материальной культуры 2-й этап изучают также методами археологии. На 3-м этапе эволюционные события происходят в основном на молекулярно-генетическом уровне и проявляются на популяционном уровне. Основными способами изучения эволюции современного человека поэтому являются биохимический, цитогенетический и популяционно-статистический методы.
Изучение первых двух этапов эволюции человека сталкивается с необходимостью датировки палеонтологического материала и элементов материальной культуры. Для определения абсолютного возраста ископаемых остатков человека и его предков широко используют физические методы, в частности радиометрические. С помощью массспектрометров определяют изотопный состав изучаемого объекта и по соотношению элементов с учетом периода полураспада входящих в его состав радиоактивных изотопов выявляют возраст образца. Ископаемый костный материал содержит в своем составе минеральные компоненты и белок коллаген, разрушающийся чрезвычайно медленно. На этом основан коллагеновый метод абсолютной датировки ископаемых остатков; чем меньше коллагена содержится в образце, тем более велика его древность.
|
|
|
В последнее время в антропологии все более активно применяют методы иммунологии, молекулярной биологии и цитогенетики. В связи с огромной важностью этих методов остановимся на них подробнее. Для определения прямого родства организмов друг с другом используют иммунологический метод, основанный на изучении иммунологических реакций антиген - антитело. Его можно применять для изучения степени родства не только современного человека с человекообразными обезьянами, но и ныне живущих видов с ископаемыми. Для этого следовые количества белка, извлекаемые из костей ископаемых форм, используют для получения антител, которые и применяют в иммунных реакциях с белками современных видов. Из современных человекообразных обезьян к человеку иммунологически наиболее близок шимпанзе, наиболее далеко от человека отстоит орангутан.
|
|
|
Иммунологическим методом было обнаружено, что белки рамапитека, человекообразной обезьяны Южной Азии (абс. возраст 13 млн. лет), более сходны с белками орангутана, чем человека и шимпанзе. Эти данные вместе с результатами морфологических и палеонтологических сопоставлений заставили отказаться от представления о том, что рамапитек является прямым предком человека, и связать его с эволюционной линией орангутана. Из этого следует, что разделение человеческой линии эволюции с африканскими человекообразными обезьянами произошло значительно позже, чем 13 млн. лет назад.
В последние годы антропогенез эффективно изучают также биомолекулярными методами. В основе этих методов изучения эволюции лежит допущение, что мера сходства двух таксонов соответствует мере их родства. Поэтому организмы, имевшие общего предка в недалеком прошлом, будут более сходными друг с другом, чем имевшие его очень давно. Сущность биомолекулярных методов состоит в использовании данных, полученных при сопоставлении белков и нуклеиновых кислот организмов разных видов для определения их родства и древности соответствующих филогенетических ветвей. При этом считают, что степень различий в аминокислотном составе белков и в нуклеотидных последовательностях ДНК позволяет судить о времени расхождения сравниваемых видов от предковой формы. Естественно, что оценивать степень родства и давность расхождения филогенетических ветвей можно лишь по накоплению нейтральных мутаций в геноме, так как вредные мутации быстро элиминируются из генофондов. В действительности нельзя считать большинство мутаций селективно нейтральными.
|
|
|
Расчеты показывают, что при нейтральности мутаций скорость их накопления постоянна только при измерении времени в количестве поколений, а не в годах. Так как продолжительность жизни поколений у различных видов различна, то и скорости накопления различий нуклеотидных последовательностей будут более велики у короткоживущих видов по сравнению с долгоживущими. Кроме того, фактическая скорость эволюции может значительно варьировать в различные временные интервалы в различных группах и по разным признакам. В определении применимости методов молекулярной биологии имеет значение возможность возникновения конвергентного сходства молекул, причем вероятность его повышается с увеличением изучаемых временных интервалов. Эволюция генов и белков часто может опережать реальное расхождение популяций, в первую очередь за счет адаптивного генетического полиморфизма. Однако молекулярно-биологические методы применимы для оценки родства и времени дивергенции в качестве приблизительных «молекулярных часов» при сравнении средних скоростей замен нуклеотидов в ДНК в целом и аминокислот во многих белках за длительные интервалы времени.
|
|
|
Биомолекулярный подход - лишь один из путей определения эволюционных расстояний, который работает только наряду с классическими методами палеонтологии и антропологии, причем в результатах при этом возможны серьезные расхождения. Так, при
изучении скелета человека, обнаруженного на территории Эквадора, по данным радиоуглеродного и аминокислотного анализа была установлена его древность в 28 тыс. лет. При использовании же коллагенового анализа возраст того же скелета оказался датированным не более чем в 2,5 тыс. лет.
|
|
В антропологии применяют несколько методов оценки степени дифференцированности таксонов, дополняющих друг друга; гибридизация ДНК, определение аминокислотных последовательностей белков, изучение генов гистосовместимости и т.д. Наиболее достоверные данные получены путем гибридизации ДНК, так как скорость эволюции ДНК в целом более постоянна, чем изменения белков. Гибридизация ДНК показала, что момент дивергенции эволюционных ветвей человека и шимпанзе наступил 6,5—6,7 млн. лет назад
Однако между эволюцией структуры генома в виде накопления генных мутаций и морфофизиологической эволюцией часто нет прямой зависимости. Это может быть связано с тем, что в формировании практически всех сложных фенотипических признаков принимают участие различные генные системы. Таким образом, скорость эволюции белков у двух разных родственных видов может быть одинакова, а скорость эволюции в целом, оцененная по комплексу фенотипических признаков, при этом оказывается различной. Сравнение аминокислотных последовательностей белков шимпанзе и человека привело к выводу, что около 99 % их белков абсолютно идентичны. Из этого следует, что и структурные гены человека и шимпанзе сходны в наибольшей степени. С чем же связаны столь значительные морфо-физиологические отличия обоих видов? Можно предполагать, что это зависит от различного распределения белков в клетках организма в процессе развития, что, в свою очередь, определяется различиями программы считывания сходной наследственной информации во времени и пространстве. Иными словами, это может быть обусловлено эволюционными преобразованиями в большей степени не структурных, а регуляторных генов.
|
|
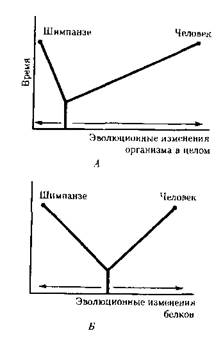
Рис. Соотношение скорости морфофизиологической (А) и
молекулярной (Б) эволюции
Поскольку изучение хромосомного материала возможно только у ныне живущих организмов, применение цитогенетического метода ограничивается современным человеком и человекообразными обезьянами. Дифференциальная окраска хромосом позволяет не только сопоставлять хромосомы разных видов приматов и человека и изучать хромосомный полиморфизм софофизиологической (А) и молекулярной временного человека, но и решать (Б) эволюции некоторые вопросы эволюции.
Выяснено, что кариотип человекообразных обезьян отличается по количеству хромосом от кариотипа человека на одну пару (23 пары хромосом человека и 24 пары шимпанзе). У человека и шимпанзе идентичны 13 пар хромосом. Хромосома 2-й пары человека точно соответствует двум соединенным с хромосомам шимпанзе, а остальные хромосомы отличаются друг от друга незначительно. Так, хромосома 5-й пары шимпанзе соответствует такой же хромосоме человека, но небольшой ее перицентрический участок инвертирован на 180° по сравнению с человеческой хромосомой. Инверсии такого рода обнаружены в кариотипах человека и шимпанзе еще в восьми хромосомах. Эти данные вместе с указаниями на сходство белков человека и шимпанзе свидетельствуют об их значительной эволюционной близости.
Сопоставление кариотипов людей, происходящих из разных популяций, приводит к выводу о полиморфизме хромосом, в первую очередь по размерам гетерохроматиновых участков. Наследуемость индивидуальных вариаций хромосом и их неравномерное распределение в разных популяциях (в частности, расовые различия по размерам длинного плеча Y-хромосомы) делают возможным популяционно-цитогенетический подход в изучении эволюции современного человека.
3.ХАРАКТЕРИСТИКА ОСНОВНЫХ ЭТАПОВ АНТРОПОГЕНЕЗА
Современные взгляды на развитие человека.
Широко распространено мнение, что человек произошел от человекообразных обезьян. Это не совсем верно.
Развитие человека и человекообразных обезьян — это не последовательные ступени, а параллельные ветви эволюции, расхождение между которыми с эволюционной точки зрения очень глубокое.
Находки палеонтологов свидетельствуют, что приматы появились около 70 млн. лет назад.
Примерно 30—40 млн. лет назад от общей исходной формы разошлись две ветви эволюции: первая, приведшая к современным высшим обезьянам, и вторая, развитие, которой в конечном счете привело к появлению человека современного анатомического типа, В дальнейшем представители этих ветвей в течение многих миллионов лет развивались независимо друг от друга. В ходе эволюции у представителей каждой из двух ветвей выработались специфические особенности биологической организации, связанные с приспособлением к соответствующему образу жизни.
Древнейшими общими предками человека и человекообразных обезьян (антропоидов) принято считать парапитеков — малоспециализированных высших обезьян, которые вели как древесный, так и наземный образ жизни. От них произошла вымершая впоследствии сборная группа обезьян — дриопитеки.
Ученые считают, что именно от какой-то группы дриопитеков, например рамапитеков, начинается эволюционная линия гоминид.
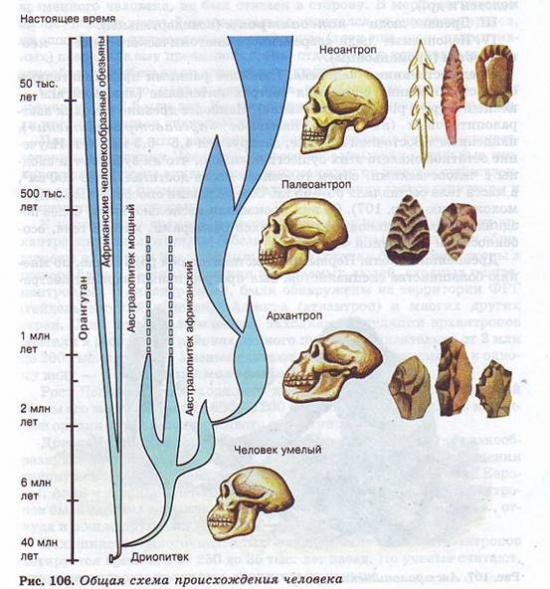
Отечественные антропологи, внесшие большой вклад в разработку теории эволюции человека, выделяют четыре стадии антропогенеза.
I. Предшественники человека — австралопитековые.
II. Древнейшие люди — прогрессивные австралопитеки, архантропы (питекантропы, синантропы, атлантропы, гейдельбергский человек и др.).
III. Древние люди — палеоантропы (неандертальцы).
IV. Ископаемые люди современного анатомического типа — неоантропы (кроманьонцы).
Предшественники человека.
Наиболее ранними представителями семейства гоминид считаются австралопитековые (лат. australis — южный и греч. pithecos — обезьяна). Наиболее древние находки австралопитековых (их иногда называют «предавстралопитеки»), найденные в Восточной Африке, датируются 4,5—6,5 млн лет. Изучение остатков скелета этих существ показало, что их зубы почти сходны с человеческими, объем головного мозга достигал более 500 см3, а масса тела составляла 35—55 кг. Они обладали способностью к прямохождению. От них произошли несколько видов более поздних австралопитеков, отличавшихся размерами, массой тела, особенностями в строении черепа.
Древнейшие люди.
Первыми представителями рода Homo, по мнению большинства специалистов, был прогрессивный вариант австралопитека — Человек умелый (Н. habilis). Это название связано с тем, что эти существа умели изготовлять примитивные режущие и рубящие орудия из гальки.

Рост Человека умелого достигал 150 см. объем мозга был на 100 см3 больше, чем у других австралопитеков и имел массу до 650 г. Он имел зубы человеческого типа, а первый палец стопы, как и у современного человека, не был отведен в сторону. В морфологическом плане Человек умелый незначительно отличался от австралопитеков, но способность изготовления орудий труда (хотя еще очень примитивных) позволила ему преодолеть грань, отделявшую ископаемых человекообразных обезьян от древнейших людей.
Считается, что первая популяция человеческих существ имеет древность 2—2,5 млн лет. Но, по мнению многих ученых, первые люди могли появиться значительно раньше.
Предполагают, что, расселяясь и попадая в новые условия существования, популяции Человека умелого образовывали отдельные изолированные формы архантропов, занимающих по многим существенным признакам промежуточное положение между австралопитеком и современным человеком.
К настоящему времени известно несколько ископаемых форм архантропов: питекантроп (обезьяночеловек), его останки были обнаружены на острове Ява; синантроп, его останки были обнаружены в пещере близ г. Пекина.
Останки ископаемых людей, сходных с питекантропами и синантропами, были обнаружены на территории ФРГ (гейдельбергский человек), Алжира (атлантроп) и многих других стран. Судя по археологическим находкам, популяции архантропов обитали в различных районах земного шара приблизительно от 2 млн до 200 тыс. лет назад. "Ученые считают, что все они относились к одному виду — Человек прямоходящий (Н. erectus).
Рост Человека прямоходящего достигал 160 см и более. Средний объем его мозга составлял 800—1200 см3. Он умел изготовлять каменные орудия труда и поддерживать огонь.

Древние люди.
Палеоантропы представляли собой крайне разнообразную как в морфологическом, так и хронологическом отношении группу ископаемых людей. Они заселяли обширные территории Европы, Азии и Африки. Первая находка скелетных останков палеоантропов была сделана в долине реки
Неандерталь в Германии в 1856 г., откуда и пошло другое их название — неандертальцы.
Большинство многочисленных находок останков палеоантропов датируется временем от 250 до 35 тыс. лет назад. Но ученые считают, что палеоантропы появились значительно раньше.
Древние люди имели объем черепной коробки около 1400 см3, изготовляли разнообразные каменные орудия (рубила, скребла, остроконечники и др.), обладали зачатками членораздельной речи
Ученые еще не пришли к общему мнению о месте и роли неандертальцев в антропогенезе и выдвинули три гипотезы:
— неандертальцы — прямые предки человека современного типа;
— неандертальцы внесли некоторый генетический вклад в генофонд человека современного типа;
— неандертальцы являются боковой тупиковой ветвью эволюции гоминидов.

Многие находки ископаемых останков в разных регионах свидетельствуют о значительной изменчивости популяций ранних неандертальцев. В настоящее время неандертальцев считают одним из подвидов Человека разумного.
Люди современного анатомического типа.
Последняя стадия эволюции человека — современные люди, или неоантропы. Впервые костные останкнеоантропов были найдены в гроте Кро Маньон во Франции в 1868 г., откуда и пошло другое название — кроманьонцы.
В целом кроманьонцы уже не имели существенных отличий от ныне живущих людей. Рост их составлял до 180 см, объем мозга — до 1600 см3. Мозговой отдел их черепа преобладал над лицевым, сплошной надглазничный валик отсутствовал, развитый подбородочный выступ указывал на то, что они смогли общаться с помощью членораздельной речи. Не окончательно решенным остается вопрос о возникновении подвида Н, sapiens sapiens, к которому принадлежит современное человечество.

Анализ палеонтологических материалов показал, что можно выделить три типа ископаемых людей, живших одновременно: неандертальцев, людей современного типа и промежуточные формы. Это дает основание предположить, что неандертальцы и кроманьонцы долгое время сосуществовали рядом и нередки были случаи смешения (метисации ). Остатки подобных промежуточных форм предков современных людей, в которых сочетались черты и неандертальцев, и кроманьонцев недавно были найдены на Ближнем Востоке на территории современного Израиля.После длительного сосуществования двух подвидов гоминид, примерно 40 тыс. лет назад, произошел демографический взрыв в популяциях людей современного анатомического типа, который сопровождался увеличением плотности населения и прогрессивными изменениями в области материальной культуры. В тяжелых условиях ледникового периода неандертальцы, очевидно,
не выдержали конкурентной борьбы с кроманьонцами, были ими вытеснены и, возможно, частично истреблены.
Дальнейшие исследования палеоантропологов и специалистов других специальностей, несомненно, внесут ясность в еще не решенные вопросы антропогенеза.
Долгое время многие палеонтологи и антропологи считали наиболее древними представителями семейства гоминид южно-африканских двуногих приматов Australopithecus africanus. Возраст большей части южно-африканских находок австралопитеков определяется в интервале 3—1 млн. лет. Строение скелетов этих организмов свидетельствует об их прямохождении. Масса мозга 450—550 г при
общей массе тела от 25 до 65 кг. Зубная система близка к человеческой: клыки малых размеров, зубы расположены в виде широкой дуги, как у человека. Это свидетельствует о всеядности.
В местах обнаружения остатков австралопитеков имеется множество костей, расколотых тяжелыми предметами Большое число черепов животных расколоты с левой стороны, и это свидетельствует о том, что австралопитеки были в основном правшами. Некото-
рые австралопитеки, видимо, начинали осваивать огонь. В 1960-1970 гг. в результате раскопок в Эфиопии и позже в Танзании и Кении были обнаружены большие группы гоминид, древность которых определяется в 4—2,8 млн. лет. По этим материалам в
1978 г. был описан новый вид австралопитеков A. afarensis, более примитивный, чем A. africanus, но тоже двуногий и с чертами «промежуточности» по отношению к современным человекообразным обезьянам и человеку (рис. 15.4). Объем мозговой полости черепа оказался в пределах 380—450 см3, т е. практически не отличался от объема мозга современного шимпанзе.
|
|

Рис. Австралопитек афарский. Реконструкция
Действительно, австалопитеки имеют большее сходство с человеком не по объему и строению мозга, а по строению зубной системы и способу передвижения. Изучение австралопитековых показало, что именно двуногость, а не большой объем мозга, явилась ключевой адаптацией ранних гоминид. В 1964 г. по находкам, сделанным
в Танзании, был выделен вид Homo habilis, имеющий абсолютный возраст 2—1,7 млн. лет. Отличительными чертами его являются двуногость, в целом прогрессивное строение кисти, зубной системы, объем мозговой коробки от 540 до 700 см3, что примерно в полтора раза превышает объем мозга австралопитеков. На внутренней поверхности чере-
па обнаруживаются признаки прогрессивных нейроморфологических изменений, определяющиеся по отпечаткам головного мозга: выраженная асимметрия полушарий и развитие двух речевых центров как условие для возникновения членораздельной речи. Большой палец стопы не отведен в сторону. Это свидетельствует о том, что морфологические перестройки, связанные с прямохождением, у него полностью завершились. Вместе с остатками Н. habilis найдены орудия труда со следами целенаправленной обработки, свидетельствующие о ранних формах трудовой деятельности.
Перечисленные признаки, ведущим из которых является прогрессивное развитие мозга, характеризуют организм уже с иной морфофункциональной организацией по сравнению с австралопитеками. Эти признаки считаются специфичными для рода Homo. Сопоставление морфологии африканского и афарского австралопитеков с Н. habilis и современным человеком позволяет предположить следующую схему филогенетических взаимоотношений ранних гоминид (рис. 15.5). Африканский австралопитек, считавшийся раньше общим предком как рода Homo, так и других австралопитековых, является в этой схеме представителем боковой ветки эволюции, приведшей к узкой специализации и образованию мощных форм типа A. robustus, которые вымерли около 1 млн. лет назад. Общим предком всех австралопитековых и рода Homo является в соответствии с этой схемой A. afarensis.
|
|
Таким образом, на протяжении 1—1,5 млн. лет представители двух близких родов и, возможно, нескольких видов семейства гоминид сосуществовали, причем не только во времени, но и на перекрывающихся территориях. В основе дивергенции различных линий ранних гоминид и австралопитековых могли лежать разного рода механизмы изоляции, в первую очередь генетические: мутации в виде хромосомных перестроек. Это означает, что эволюция австралопитековых шла постепенно, приводя благодаря дивергенции к морфологическому и экологическому разнообразию.
Дата добавления: 2021-05-18; просмотров: 301; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!