Экологические группы растений по отношению к свету и их адаптивные особенности
Экологические группы растений по отношению к воде
Гидатофиты – это водные растения, целиком или почти целиком погруженные в воду. Среди них – цветковые, которые вторично перешли к водному образу жизни (элодея, рдесты, водяные лютики, валлиснерия, уруть и др.). Вынутые из воды, эти растения быстро высыхают и погибают. У них редуцированы устьица и нет кутикулы. Транспирация у таких растений отсутствует, а вода выделяется через особые клетки – гидатоды.
Листовые пластинки у гидатофитов, как правило, тонкие, без дифференцировки мезофилла, часто рассеченные, что способствует более полному использованию ослабленного в воде солнечного света и усвоению СО2. Нередко выражена разнолистность – гетерофиллия; у многих видов есть плавающие листья, имеющие световую структуру. Поддерживаемые водой побеги часто не имеют механических тканей, в них хорошо развита аэренхима (рис. 32).
Корневая система цветковых гидатофитов сильно редуцирована, иногда отсутствует совсем или утратила свои основные функции (у рясок). Поглощение воды и минеральных солей происходит всей поверхностью тела. Цветоносные побеги, как правило, выносят цветки над водой (реже опыление совершается в воде), а после опыления побеги снова могут погружаться, и созревание плодов происходит под водой (валлиснерия, элодея, рдесты и др.).
Гидрофиты – это растения наземно-водные, частично погруженные в воду, растущие по берегам водоемов, на мелководьях, на болотах. Встречаются в районах с самыми разными климатическими условиями. К ним можно отнести тростник обыкновенный, частуху подорожниковую, вахту трехлистную, калужницу болотную и другие виды. У них лучше, чем у гидатофитов, развиты проводящие и механические ткани. Хорошо выражена аэренхима. В аридных районах при сильной инсоляции их листья имеют световую структуру. У гидрофитов есть эпидерма с устьицами, интенсивность транспирации очень высока, и они могут расти только при постоянном интенсивном поглощении воды.
|
|
|
Гигрофиты – наземные растения, живущие в условиях повышенной влажности воздуха и часто на влажных почвах. Среди них различают теневые и световые. Теневые гигрофиты – это растения нижних ярусов сырых лесов в разных климатических зонах (недотрога, цирцея альпийская, бодяк огородный, многие тропические травы и т. п.). Из-за высокой влажности воздуха у них может быть затруднена транспирация, поэтому для улучшения водного обмена на листьях развиваются гидатоды, или водяные устьица, выделяющие капельно-жидкую воду. Листья часто тонкие, с теневой структурой, со слабо развитой кутикулой, содержат много свободной и малосвязанной воды. Обводненность тканей достигает 80 % и более. При наступлении даже непродолжительной и несильной засухи в тканях создается отрицательный водный баланс, растения завядают и могут погибнуть.
|
|
|
К световым гигрофитам относятся виды открытых местообитаний, растущие на постоянно влажных почвах и во влажном воздухе (папирус, рис, сердечники, подмаренник болотный, росянка и др.). Переходные группы – мезогигрофиты и гигромезофиты.
Мезофиты могут переносить непродолжительную и не очень сильную засуху. Это растения, произрастающие при среднем увлажнении, умеренно теплом режиме и достаточно хорошей обеспеченности минеральным питанием. К мезофитам можно отнести вечнозеленые деревья верхних ярусов тропических лесов, листопадные деревья саванн, древесные породы влажных вечнозеленых субтропических лесов, летнезеленые лиственные породы лесов умеренного пояса, кустарники подлеска, травянистые растения дубравного широкотравья, растения заливных и не слишком сухих суходольных лугов, пустынные эфемеры и эфемероиды, многие сорные и большинство культурных растений. Из приведенного перечня видно, что группа мезофитов очень обширна и неоднородна. По способности регулировать свой водный обмен одни приближаются к гигрофитам (мезогигрофиты), другие – к засухоустойчивым формам (мезоксерофиты).
|
|
|
Ксерофиты растут в местах с недостаточным увлажнением и имеют приспособления, позволяющие добывать воду при ее недостатке, ограничивать испарение воды или запасать ее на время засухи. Ксерофиты лучше, чем все другие растения, способны регулировать водный обмен, поэтому и во время продолжительной засухи остаются в активном состоянии. Это растения пустынь, степей, жестколистных вечнозеленых лесов и кустарниковых зарослей, песчаных дюн.
Ксерофиты подразделяются на два основных типа: суккуленты и склерофиты.
Суккуленты– сочные растения с сильно развитой водозапасающей паренхимой в разных органах. Стеблевые суккуленты – кактусы, стапелии, кактусовидные молочаи; листовые суккуленты – алоэ, агавы, мезембриантемумы, молодило, очитки; корневые суккуленты – аспарагус. В пустынях Центральной Америки и Южной Африки суккуленты могут определять облик ландшафта (рис. 33).
Листья, а в случае их редукции стебли суккулентов имеют толстую кутикулу, часто мощный восковой налет или густое опушение. Устьица погруженные, открываются в щель, где задерживаются водяные пары.
|
|
|
Днем они закрыты. Это помогает суккулентам сберегать накопленную влагу, но зато ухудшает газообмен, затрудняет поступление СО2 внутрь растения. Поэтому многие суккуленты из семейств лилейных, бромелиевых, кактусовых, толстянковых ночью при открытых устьицах поглощают СО2, который только на следующий день перерабатывают в процессе фотосинтеза. Поглощенный СО2 переводится в малат. Кроме того, при дыхании ночью углеводы разлагаются не до углекислого газа, а до органических кислот, которые отводятся в клеточный сок. Днем на свету малат и другие органические кислоты расщепляются с выделением СО2, который используется в процессе фотосинтеза. Таким образом, крупные вакуоли с клеточным соком запасают не только воду, но и СО2. Так как у суккулентов ночная фиксация углекислоты и переработка ее днем в ходе фотосинтеза разделены во времени, они обеспечивают себя углеродом, не подвергаясь риску чрезмерной потери воды, но масштабы поступления углекислого газа при таком способе невелики, и растут суккуленты медленно.
Осмотическое давление клеточного сока суккулентов мало – всего 3 · 105 – 8 · 105 Па (3–8 атм), они развивают небольшую сосущую силу и способны всасывать воду лишь атмосферных осадков, просочившихся в верхний слой почвы. Корневая система их неглубокая, но сильно распростертая, что особенно характерно для кактусов.
Склерофиты– это растения, наоборот, сухие на вид, часто с узкими и мелкими листьями, иногда свернутыми в трубочку. Листья могут быть также рассеченными, покрытыми волосками или восковым налетом. Хорошо развита склеренхима, поэтому растения без вредных последствий могут терять до 25 % влаги не завядая. В клетках преобладает связанная вода. Сосущая сила корней до нескольких десятков атмосфер, что позволяет успешно добывать воду из почвы. При недостатке воды резко снижают транспирацию. Склерофиты можно подразделить на две группы: эуксерофитов и стипаксерофитов.
К эуксерофитам относятся многие степные растения с розеточными и полурозеточными, сильно опушенными побегами, полукустарнички, некоторые злаки, полынь холодная, эдельвейс эдельвейсовидный и др. Наибольшую биомассу эти растения создают в период, благоприятный для вегетации, а в жару уровень обменных процессов у них очень низок.
Стипаксерофиты– это группа узколистных дерновинных злаков (ковыли, тонконоги, типчак и др.). Характеризуются низкой транспирацией в засушливый период и могут переносить особенно сильное обезвоживание тканей. Свернутые в трубочку листья имеют внутри влажную камеру. Транспирация идет через погруженные в бороздки устьица внутрь этой камеры, что снижает потери влаги (рис. 34).
Кроме названных экологических групп растений, выделяют еще целый ряд смешанных или промежуточных типов.
Различные пути регуляции водообмена позволили растениям заселить самые различные по экологическим условиям участки суши. Многообразие приспособлений лежит, таким образом, в основе распространения растений по поверхности земли, где дефицит влаги является одной из главных проблем экологических адаптаций.
Экологические группы растений по отношению к свету и их адаптивные особенности
Световой режим любого местообитания определяется интенсивностью прямого и рассеянного света, количеством света (годовой суммарной радиацией), его спектральным составом, а также альбедо – отражательной способностью поверхности, на которую падает свет.
Перечисленные элементы светового режима очень переменчивы и зависят от географического положения, высоты над уровнем моря, от рельефа, состояния атмосферы, характера земной поверхности, растительности, от времени суток, сезона года, солнечной активности и глобальных изменений в атмосфере.
У растений возникают различные морфологические и физиологические адаптации к световому режиму местообитаний.
По требованию к условиям освещения принято делить растения на следующие экологические группы:
1) светолюбивые (световые), или гелиофиты, – растения открытых, постоянно хорошо освещаемых местообитаний;
2) тенелюбивые (теневые), или сциофиты, – растения нижних ярусов тенистых лесов, пещер и глубоководные растения; они плохо переносят сильное освещение прямыми солнечными лучами;
3) теневыносливые, или факультативные гелиофиты, – могут переносить большее или меньшее затенение, но хорошо растут и на свету; они легче других растений перестраиваются под влиянием изменяющихся условий освещения.
Можно отметить некоторые общие приспособительные особенности, свойственные растениям каждой экологической группы.
Световые адаптации гелиофитов и сциофитов. Гелиофиты часто имеют побеги с укороченными междоузлиями, сильно ветвящиеся, нередко розеточные. Листья гелиофитов обычно мелкие или с рассеченной листовой пластинкой, с толстой наружной стенкой клеток эпидермы, нередко с восковым налетом или густым опушением, с большим числом устьиц на единицу площади, часто погруженных, с густой сетью жилок, с хорошо развитыми механическими тканями. У ряда растений листья фотометричные, т. е. повернуты ребром к полуденным лучам или могут менять положение своих частей в зависимости от высоты стояния Солнца. Так, у степного растения софоры листочки непарноперистосложного листа в жаркий день подняты вверх и сложены, у василька русского так же ведут себя сегменты перисторассеченного листа.
Оптический аппарат гелиофитов развит лучше, чем у сциофитов, имеет большую фотоактивную поверхность и приспособлен к более полному поглощению света. Обычно у них лист толще, клетки эпидермы и мезофилла мельче, палисадная паренхима двухслойная или многослойная (у некоторых саван-ных растений Западной Африки – до 10 слоев), нередко развита под верхней и нижней эпидермой. Мелкие хлоропласты с хорошо развитой гранальной структурой в большом числе (до 200 и более) расположены вдоль продольных стенок.
Хлорофилла на сухую массу в листьях гелиофитов приходится меньше, но зато в них содержится больше пигментов I пигментной системы и хлорофилла П700. Отношение хлорофилла а к хлорофиллу b равно примерно 5: 1. Отсюда высокая фотосинтетическая способность гелиофитов. Компенсационная точка лежит в области более высокой освещенности. Интенсивность фотосинтеза достигает максимума при полном солнечном освещении. У особой группы растений – гелиофитов, у которых фиксация СО2 идет путем С4-дикарбоновых кислот, световое насыщение фотосинтеза не достигается даже при самой сильной освещенности. Это растения из засушливых областей (пустынь, саванн). Особенно много С4-растений среди семейств мятликовых, осоковых, аизовых, портулаковых, амарантовых, маревых, гвоздичных, молочайных. Они способны к вторичной фиксации и реутилизации СО2, освобождающегося при световом дыхании, и могут фотосинтезировать при высоких температурах и при закрытых устьицах, что часто наблюдается в жаркие часы дня.
Обычно С4-растения, особенно сахарный тростник и кукуруза, отличаются высокой продуктивностью.
Сциофиты– это растения, постоянно находящиеся в условиях сильного затенения. При освещенности 0,1–0,2 % могут расти только мхи и селягинеллы. Плауны довольствуются 0,25-0,5 % полного дневного света, а цветковые растения встречаются обычно там, где освещенность в пасмурные дни достигает не менее 0,5–1% (бегонии, недотрога, травы из семейств имбирные, мареновые, коммелиновые).
В северных широколиственных и темнохвойных лесах полог сомкнутого древостоя может пропускать всего 1–2% ФАР, изменяя ее спектральный состав. Сильнее всего поглощаются синие и красные лучи, и пропускается относительно больше желто-зеленых лучей, дальних красных и инфракрасных. Слабая освещенность сочетается с повышенной влажностью воздуха и повышенным содержанием в нем СО2, особенно у поверхности почвы. Сциофиты этих лесов – зеленые мхи, плауны, кислица обыкновенная, грушанки, майник двулистный и др.
Листья у сциофитов располагаются горизонтально, нередко хорошо выражена листовая мозаика. Листья темно-зеленые, более крупные и тонкие. Клетки эпидермы крупнее, но с более тонкими наружными стенками и тонкой кутикулой, часто содержат хлоропласты. Клетки мезофилла крупнее, палисадная паренхима однослойная или имеет нетипичное строение и состоит не из цилиндрических, а из трапециевидных клеток. Площадь жилок вдвое меньше, чем у листьев гелиофитов, число устьиц на единицу площади меньше. Хлоропласты крупные, но число их в клетках невелико.
У сциофитов по сравнению с гелиофитами меньше хлорофилла П700. Отношение хлорофилла а к хлорофиллу b равно примерно 3: 2. С меньшей интенсивностью протекают у них такие физиологические процессы, как транспирация, дыхание. Интенсивность фотосинтеза, быстро достигнув максимума, перестает возрастать при усилении освещенности, а на очень ярком свету может даже понизиться.
У лиственных теневыносливых древесных пород и кустарников (дуба черешчатого, липы сердцевидной, сирени обыкновенной и др.) листья, расположенные по периферии кроны, имеют структуру, сходную со структурой листьев гелиофитов, и называются световыми, а в глубине кроны – теневые листья с теневой структурой, сходной со структурой листьев сциофитов (рис. 24).
Факультативные гелиофиты, или теневыносливые растения, в зависимости от степени теневыносливости имеют приспособительные особенности, сближающие их то с гелиофитами, то со сциофитами. К этой группе можно отнести некоторые луговые растения, лесные травы и кустарники, растущие и в затененных участках леса, и на лесных полянах, опушках, вырубках. На осветленных местах они разрастаются часто сильнее, однако оптимальное использование ФАР у них происходит не при полном солнечном освещении.
У деревьев и кустарников теневая или световая структура листа часто определяется условиями освещения предыдущего года, когда закладываются почки: если закладка почек идет на свету, то формируется световая структура, и наоборот.
Если в одном и том же местообитании закономерно периодически изменяется световой режим, растения в разные сезоны могут проявлять себя то как светолюбивые, то как теневыносливые.
Весной в дубравах под полог леса проникает 50–60 % солнечной радиации. Листья розеточных побегов сныти обыкновенной имеют световую структуру и отличаются высокой интенсивностью фотосинтеза. В это время они создают основную часть органического вещества годичной продукции. Листья сныти летней генерации, появляющиеся при развитом древесном пологе, под который проникает в среднем 3,5 % солнечной радиации, имеют типичную теневую структуру, и интенсивность фотосинтеза их значительно ниже, в 10–20 раз. Подобную двойственность по отношению к свету проявляет и осока волосистая, светолюбивая весной и теневыносливая летом. По-видимому, это свойственно и другим растениям дубравного широкотравья.
Отношение к световому режиму меняется у растений и в онтогенезе. Проростки и ювенильные растения многих луговых видов и древесных пород более теневыносливы, чем взрослые особи.
Иногда у растений меняются требования к световому режиму, когда они оказываются в иных климатических и эдафических условиях. Так, обычные теневыносливые растения хвойного леса – черника, седмичник европейский и некоторые другие – в тундре приобретают особенности гелиофитов.
Наиболее общая адаптация растений к максимальному использованию ФАР – пространственная ориентация листьев. При вертикальном расположении листьев, как, например, у многих злаков и осок, солнечный свет полнее поглощается в утренние и вечерние часы – при более низком стоянии солнца. При горизонтальной ориентации листьев полнее используются лучи полуденного солнца. При диффузном расположении листьев в разных плоскостях солнечная радиация в течение дня утилизируется наиболее полно. Обычно при этом листья нижнего яруса на побеге отклонены горизонтально, среднего направлены косо вверх, а верхнего располагаются почти вертикально.
Считают, что кукуруза является одной из самых высокопродуктивных сельскохозяйственных культур потому, что наряду с высоким КПД фотосинтеза у нее наблюдается диффузное расположение листьев, при котором полнее поглощается ФАР.
На севере, где высота стояния солнца меньше, встречается больше растений с вертикальным расположением листьев, а на юге – с горизонтальным. Для получения большей биомассы выгодны также посевы и насаждения, в которых сочетаются растения с разной пространственной ориентацией листьев, причем в верхнем ярусе лучше иметь растения с вертикальным расположением листьев, которые полнее используют свет при низком стоянии солнца, не препятствуют прохождению полуденных лучей к расположенным в нижнем ярусе листьям с горизонтальной ориентацией.
Обитатели почвы
Неоднородность почвы приводит к тому, что для организмов разных размеров она выступает как разная среда. Для микроорганизмов особое значение имеет огромная суммарная поверхность почвенных частиц, так как на них адсорбируется подавляющая часть микробного населения. Сложность почвенной среды создает большое разнообразие условий для самых разных функциональных групп: аэробов и анаэробов, потребителей органических и минеральных соединений. Для распределения микроорганизмов в почве характерна мелкая очаговость, поскольку даже на протяжении нескольких миллиметров могут сменяться разные экологические зоны.
Для мелких почвенных животных (рис. 52, 53), которых объединяют под названием микрофауна (простейшие, коловратки, тихоходки, нематоды и др.), почва – это система микроводоемов. По существу, это водные организмы. Они живут в почвенных порах, заполненных гравитационной или капиллярной водой, а часть жизни могут, как и микроорганизмы, находиться в адсорбированном состоянии на поверхности частиц в тонких прослойках пленочной влаги. Многие из этих видов обитают и в обычных водоемах. Однако почвенные формы намного мельче пресноводных и, кроме того, отличаются способностью долго находиться в инцистированном состоянии, пережидая неблагоприятные периоды. В то время как пресноводные амебы имеют размеры 50-100 мкм, почвенные – всего 10–15. Особенно мелки представители жгутиковых, нередко всего 2–5 мкм. Почвенные инфузории также имеют карликовые размеры и к тому же могут сильно менять форму тела.
Для дышащих воздухом несколько более крупных животных почва предстает как система мелких пещер. Таких животных объединяют под названием мезофауна (рис. 54). Размеры представителей мезофауны почв – от десятых долей до 2–3 мм. К этой группе относятся в основном членистоногие: многочисленные группы клещей, первичнобескрылые насекомые (коллемболы, протуры, двухвостки), мелкие виды крылатых насекомых, многоножки симфилы и др. У них нет специальных приспособлений к рытью. Они ползают по стенкам почвенных полостей при помощи конечностей или червеобразно извиваясь. Насыщенный водяными парами почвенный воздух позволяет дышать через покровы. Многие виды не имеют трахейной системы. Такие животные очень чувствительны к высыханию. Основным средством спасения от колебания влажности воздуха для них является передвижение вглубь. Но возможность миграции по почвенным полостям вглубь ограничивается быстрым уменьшением диаметра пор, поэтому передвижения по скважинам почвы доступны только самым мелким видам. Более крупные представители мезофауны обладают некоторыми приспособлениями, позволяющими переносить временное снижение влажности почвенного воздуха: защитными чешуйками на теле, частичной непроницаемостью покровов, сплошным толстостенным панцирем с эпикутикулой в сочетании с примитивной трахейной системой, обеспечивающей дыхание.
Периоды затопления почвы водой представители мезофауны переживают в пузырьках воздуха. Воздух задерживается вокруг тела животных благодаря их несмачивающимся покровам, снабженным к тому же волосками, чешуйками и т. п. Пузырек воздуха служит для мелкого животного своеобразной «физической жаброй». Дыхание осуществляется за счет кислорода, диффундирующего в воздушную прослойку из окружающей воды.
Представители микро– и мезофауны способны переносить зимнее промерзание почвы, так как большинство видов не может уходить вниз из слоев, подвергающихся воздействию отрицательных температур.
Более крупных почвенных животных, с размерами тела от 2 до 20 мм, называют представителями макрофауны (рис. 55). Это личинки насекомых, многоножки, энхитреиды, дождевые черви и др. Для них почва – плотная среда, оказывающая значительное механическое сопротивление при движении. Эти относительно крупные формы передвигаются в почве либо расширяя естественные скважины путем раздвигания почвенных частиц, либо роя новые ходы. Оба способа передвижения накладывают отпечаток на внешнее строение животных.
Возможность двигаться по тонким скважинам, почти не прибегая к рытью, присуща только видам, которые имеют тело с малым поперечным сечением, способное сильно изгибаться в извилистых ходах (многоножки – костянки и геофилы). Раздвигая частицы почвы за счет давления стенок тела, передвигаются дождевые черви, личинки комаров-долгоножек и др. Зафиксировав задний конец, они утончают и удлиняют передний, проникая в узкие почвенные щели, затем закрепляют переднюю часть тела и увеличивают его диаметр. При этом в расширенном участке за счет работы мышц создается сильное гидравлическое давление несжимающейся внутриполостной жидкости: у червей – содержимого целомических мешочков, а у типулид – гемолимфы. Давление передается через стенки тела на почву, и таким образом животное расширяет скважину. При этом сзади остается открытый ход, что грозит увеличением испарения и преследованием хищников. У многих видов развиты приспособления к экологически более выгодному типу передвижения в почве – рытью с закупориванием за собой хода. Рытье осуществляется разрыхлением и отгребанием почвенных частиц. Личинки разных насекомых используют для этого передний конец головы, мандибулы и передние конечности, расширенные и укрепленные толстым слоем хитина, шипами и выростами. На заднем конце тела развиваются приспособления для прочной фиксации – выдвигающиеся подпорки, зубцы, крючья. Для закрывания хода на последних сегментах у ряда видов имеется специальная вдавленная площадка, обрамленная хитиновыми бортиками или зубцами, своего рода тачка. Подобные площадки образуются на задней части надкрылий и у жуков-короедов, которые тоже используют их для закупоривания ходов буровой мукой. Закрывая за собой ход, животные – обитатели почвы постоянно находятся в замкнутой камере, насыщенной испарениями собственного тела.
Газообмен большинства видов этой экологической группы осуществляется при помощи специализированных органов дыхания, но наряду с этим дополняется газообменом через покровы. Возможно даже исключительно кожное дыхание, например у дождевых червей, энхитреид.
Роющие животные могут уходить из слоев, где возникает неблагоприятная обстановка. В засуху и к зиме они концентрируются в более глубоких слоях, обычно в нескольких десятках сантиметров от поверхности.
Мегафауна почв – это крупные землерои, в основном из числа млекопитающих. Ряд видов проводит в почве всю жизнь (слепыши, слепушонки, цокоры, кроты Евразии, златокроты
Африки, сумчатые кроты Австралии и др.). Они прокладывают в почве целые системы ходов и нор. Внешний облик и анатомические особенности этих животных отражают их приспособленность к роющему подземному образу жизни. У них недоразвиты глаза, компактное, вальковатое тело с короткой шеей, короткий густой мех, сильные копательные конечности с крепкими когтями. Слепыши и слепушонки разрыхляют землю резцами. К мегафауне почвы следует отнести и крупных олигохет, в особенности представителей семейства Megascolecidae, обитающих в тропиках и Южном полушарии. Самый крупный из них австралийский Megascolides australis достигает в длину 2,5 и даже 3 м.
Кроме постоянных обитателей почвы, среди крупных животных можно выделить большую экологическую группу обитателей нор (суслики, сурки, тушканчики, кролики, барсуки и т. п.). Они кормятся на поверхности, но размножаются, зимуют, отдыхают, спасаются от опасности в почве. Целый ряд других животных использует их норы, находя в них благоприятный микроклимат и укрытие от врагов. Норники обладают чертами строения, характерными для наземных животных, но имеют ряд приспособлений, связанных с роющим образом жизни. Например, у барсуков длинные когти и сильная мускулатура на передних конечностях, узкая голова, небольшие ушные раковины. У кроликов по сравнению с зайцами, не роющими нор, заметно укорочены уши и задние ноги, более прочный череп, сильнее развиты кости и мускулатура предплечий и т. п.
По целому ряду экологических особенностей почва является средой, промежуточной между водной и наземной. С водной средой почву сближают ее температурный режим, пониженное содержание кислорода в почвенном воздухе, насыщенность его водяными парами и наличие воды в других формах, присутствие солей и органических веществ в почвенных растворах, возможность двигаться в трех измерениях.
С воздушной средой почву сближают наличие почвенного воздуха, угроза иссушения в верхних горизонтах, довольно резкие изменения температурного режима поверхностных слоев.
Промежуточные экологические свойства почвы как среды обитания животных позволяют предполагать, что почва играла особую роль в эволюции животного мира. Для многих групп, в частности членистоногих, почва послужила средой, через которую первоначально водные обитатели смогли перейти к наземному образу жизни и завоевать сушу. Этот путь эволюции членистоногих доказан трудами М. С. Гилярова
Датский ботаник К. Раункиер рассматривал жизненные формы как результат приспособления растений к внешней среде. Решающее значение он придавал климату. В основу выделения жизненных форм Раункиер положил различия в приспособлении растений к переживанию неблагоприятного времени года. Эта приспособленность отражена в размещении почек или верхушек побегов по отношению к поверхности почвы (рис. 69).
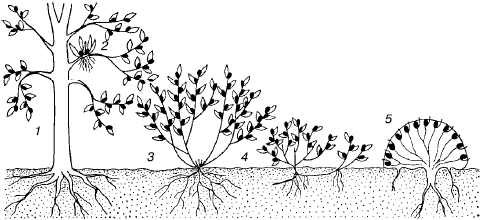
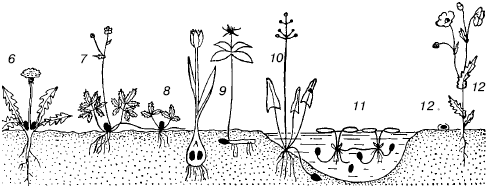
Рис. 69. Жизненные формы растений (по К. Раункиеру, 1907):
1–3– фанерофиты; 4–5 – хамефиты; 6–7 – гемикриптофиты; 8-11 – криптофиты; 12 – терофиты; 12а – семя с зародышем
Система К. Раункиера
К. Раункиер использовал для классификации жизненных форм растений единственный, но имеющий большое приспособительное значение признак – положение почек возобновления по отношению к поверхности почвы. Сначала он разработал эту систему для растений Средней Европы, но затем распространил на растения всех климатических поясов.
Все растения Раункиер разделил на пять типов (1903), в которых позднее выделил подтипы (1907).
1. Фанерофиты. Почки возобновления или верхушки побегов расположены в течение неблагоприятного времени года более или менее высоко в воздухе и подвергаются всем превратностям погоды. Подразделяются на 15 подтипов по высоте растений, по ритму развития листвы, по степени защищенности почек, по консистенции стебля. Один из подтипов – эпифитные фанерофиты.
2. Хамефиты. Почки возобновления у поверхности почвы или не выше 20–30 см. Зимой прикрыты снежным покровом. Подразделяются на 4 подтипа.
3. Гемикриптофиты. Почки возобновления или верхушки побегов на поверхности почвы, часто прикрыты подстилкой. Включают три подтипа и более мелкие подразделения.
4. Криптофиты. Почки возобновления или верхушки побегов сохраняются в почве (геофиты) или под водой (гелофиты и гидрофиты). Подразделяются на 7 подтипов.
5. Терофиты. Переносят неблагоприятное время года только в семенах.
Раункиер считал, что жизненные формы складываются исторически как результат приспособления растений к климатическим условиям. Процентное распределение видов по жизненным формам в растительных сообществах на изучаемой территории он назвал биологическим спектром. Для разных зон и стран были составлены биологические спектры, которые могли служить индикаторами климата. Так, жаркий и влажный климат тропиков был назван «климатом фанерофитов», умеренно-холодные области имеют «климат гемикриптофитов», полярные страны – «климат хамефитов».
Критики воззрений Раункиера отмечают, что его типы жизненных форм слишком обширны и неоднородны: хамефиты включают растения с разным отношением к климату, их много как в тундрах, так и в полупустынях. И не только современный климат определяет спектр жизненных форм, но и комплекс почвенно-литологических условий, а также история формирования флоры и влияние человеческой культуры. Тем не менее классификация жизненных форм растений по Раункиеру остается популярной и продолжает модифицироваться.
Классификации жизненных форм животных, как и растений, весьма разнообразны и зависят от принципов, которые кладут в их основу. На морфологии млекопитающих, по-видимому, в наибольшей степени сказывается характер их передвижения в разных средах. А. Н. Формозов выделил на этом основании среди зверей следующие адаптивные типы: 1) наземные формы; 2) подземные (землерои); 3) древесные; 4) воздушные и 5) водные. Между типами есть переходы.
В пределах каждой группы особенности поступательного движения и образа жизни формируют более специфические приспособительные формы. Так, наземные млекопитающие передвигаются в основном посредством ходьбы, бега и прыжков, что проявляется в их внешнем облике. Например, прыгающие животные: кенгуру, тушканчики, прыгунчики, кенгуровые крысы – отличаются компактным телом с удлиненными задними конечностями и значительно укороченными передними. Очень сильно развиты мышцы – разгибатели спины, увеличивающие мощность толчка. Длинный хвост играет роль балансира и руля, позволяющего резко изменять направление движения. Он служит также дополнительной точкой опоры. Сходный тип передвижения и внешний облик имели и некоторые вымершие динозавры мезозоя. Естественно, что, кроме общих адаптивных признаков, каждый вид отличается деталями внешнего строения в зависимости от экологической специфики.
Дата добавления: 2021-04-24; просмотров: 86; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!