Показатели содержания лимфоцитов у здоровых людей
КЛЕТОЧНЫЙ ИММУНИТЕТ.
ТИПЫ КЛЕТОЧНОЙ ЦИТОТОКСИЧНОСТИ.
РЕЦЕПТОРЫ И МАРКЕРЫ, СУБПОПУЛЯЦИИ ЛИМФОЦИТОВ.
Материал для самостоятельного освоения
Список сокращений
ГКГС – главный комплекс гистосовместимости;
ИЛ1 – ИЛ18 - интерлейкины 1-18;
ТКР - Т-клеточный рецептор ( см. англ. TcR - T-cell receptor);
CD – кластеры дифференцировки;
CTL – цитотоксические Т-лимфоциты (синоним: Т-лимфоциты эффекторы);
FcγR – рецептор для Fc фрагмента иммуноглобулина G;
HLA – (англ. Human Leukocyte Antigens) человеческий лейкоцитарный антиген;
IgG - иммуноглобулин G;
NK – натуральные киллеры
TcR – (англ. T-cell receptor) Т-клеточный рецептор;
Th1 – Т–хелперы первого типа;
Th2 – Т-хелперы второго типа;
КЛЕТОЧНЫЙ ИММУНИТЕТ
Иммунная система млекопитающих обеспечивает защиту организма двумя основными специфическими способами. Во-первых, это образование специфических антител, а во-вторых, образование и функционирование клеточных факторов приобретенного иммунитета, не только оказывающих ЭФФЕКТОРНОЕ ДЕЙСТВИЕ (разрушение клеток мишеней: опухолевых, мутировавших, и др.), но и осуществляющих РЕГУЛЯЦИЮ иммунного ответа. А так же участвующих в формировании иммунологической памяти, распознавании антигена и индукции иммунного ответа. Клетками, выполняющими такие многообразные функции, являются в первую очередь Т‑ЛИМФОЦИТЫ. Причем среди Т‑лимфоцитов существуют субпопуляции, основные из которых Т‑хелперы и Т‑эффекторы (цитотоксические Т-лимфоциты). Приобретение ими специфических функций происходит в центральном органе иммунной системы — тимусе.
|
|
|
Важной особенностью Т-лимфоцитов является способность распознавать только антигены презентированные (представленные) на поверхности вспомогательных антигенпредставляющих клеток (дендритных, макрофагов или В-лимфоцитов) в комплексе с собственными антигенами гистосовместимости.
ТИМУС важный лимфопоэтический и эндокринный орган. Главная функция тимуса это регулирующее влияние на Т-клеточный иммуногенез. Гемопоэтические стволовые клетки (предшественники Т-лимфоцитов) из костного мозга через кровяное русло мигрируют в тимус. Установлено, что гормоны тимуса, а точнее растворимые факторы тимуса, создают гуморальное микроокружение тимическим лимфоцитам (последние называют тимоцитами). Значительную роль на внутритимическое развитие Т-лимфоцитов оказывают эпителиальные клетки тимуса и дендритные клетки тимуса. Межклеточные взаимодействия тимоцитов с ними обеспечивают созревание и селекцию Т-клеток. Влияние гормонов тимуса на иммуногенез происходит не только в процессе внутритимической дифференцировки лимфоцитов, но и на расстоянии, поступая в кровь, они оказывают влияние на предшественников лимфоцитов в костном мозге и циркулирующие лимфоциты. В тимусе происходят основные начальные этапы дифференцировки Т‑лимфоцитов (тимоцитов), с последующим поступлением зрелых лимфоцитов в кровь. Среди этих лимфоцитов не должно быть аутореактивных – способных взаимодействовать с аутологичными антигенами организма индивидуума.
|
|
|
Необходимо отметить, что до сих пор окончательно не установлена биологическая роль гормональных веществ тимуса. Их эффекты не ограничиваются лимфопоэзом, они оказывают влияние на метаболизм кальция и фосфора, обмен и утилизацию глюкозы, мышечный тонус, рост и половое созревание, обладают обезболивающим действием, влияют на пигментный обмен.
Дифференцировка Т-лимфоцитов.
В процессе дифференцировки Т‑лимфоцитов выделяют два основных этапа (как Вы помните, такие же два этапа выделяют в процессе дифференцировки В-лимфоцитов):
1. Антигеннезависимая дифференцировка – происходит постоянно в тимусе.
2. Антигензависимая дифференцировка – происходит в периферических органах иммунной системы только при контакте Т-лимфоцита с антигеном.
|
|
|
АНТИГЕННЕЗАВИСИМАЯ ДИФФЕРЕНЦИРОВКА Т-ЛИМФОЦИТОВ
Родоначальной клеткой Т-лимфоцитов, как и всех клеток крови, является полипотентная стволовая гемопоэтическая клетка. Её маркером является CD 34. Справочную информацию о CD смотри в конце учебно-методической рекомендации.


|
|
|
|
|
|
По своему строению молекулы TcR (ТКР) напоминают иммуноглобулины (Fab-фрагмент) и состоят из альфа- и бета- цепей (TcR αβ их подавляющее большинство) или гамма- и дельта- цепей ( TcR γδ). αβ- и γδ – формы TcR весьма сходны по структуре. Каждая цепь ТКР состоит из двух областей (доменов): наружный вариабельный (V) , второй – константный (С). Отдельные гены кодирующие всю вариабельную область (V) α и β цепей TcR отсутствуют. Фрагменты вариабельных доменов кодируются тремя группами генов обозначаемых V, D, J. В клеточном геноме гены, кодирующие V-, J- и D-сегменты вариабельной области, представлены в виде многочисленных вариантов. Именно различные сочетания V-, J- и D-сегментов V области, образующиеся в процессе генной перестройки, называемой реаранжировкой, обеспечивают разнообразие молекул ТКР. Таким образом, ограниченное число генов (около 400) может кодировать рецепторы для почти бесконечного числа антигенов (многих миллионов). Причем различные комбинации генов V, D, J –сегментов - это только один из способов достижения многообразия антигенных рецепторов Т-лимфоцитов.
|
Основная функция зрелых Т-лимфоцитов – распознавание чужеродных антигенных пептидов в комплексе с собственными антигенами главного комплекса гистосовместимости (ГКГС) на поверхности антигенпрезентирующих клеток или на поверхности любых клеток-мишеней организма. Для выполнения этой функции Т-лимфоциты должны быть способны распознавать собственные антигены ГКГС. В то же время, Т-клетки не должны распознавать аутоантигены самого организма, связанные с собственными антигенами ГКГС. В связи с этим в тимусе молодые тимоциты проходят селекцию («отбор»), TcR которых соответствует вышеуказанным условиям.
Суть позитивной и негативной селекции состоит в следующем (см. рисунок на титульном листе):
Позитивная селекция. Т-лимфоциты, ТКР которых обладает способностью распознавать HLA (молекулы ГКГС) стромальных клеток тимуса, выживают, а если нет – то гибнут путем апоптоза. Позитивная селекция – поддержка избирательной выживаемости. Таким образом, выживают только лимфоциты способные распознавать собственные HLA! И эта способность в последующем является важной в функционировании Т-клеток.

Кроме этого в тимусе погибают путем апоптоза аутореактивные лимфоциты (лимфоциты имеющие ТКР к антигенным детерминантам собственных тканей). Важно, что при контакте с эпителиоидными клетками тимуса Т-лимфоциты, реагирующие на «своё», разрушаются путем запуска апоптоза (запрограммированной клеточной смерти при активации через CD95 – Fas рецептор). Это отрицательная селекция. В итоге, исчезают аутореактивные клоны клеток и возникает толерантность (неотвечаемость) к «своему». В тимусе около 95 – 97% лимфоцитов погибают в результате процесса селекции.
В последующем одна из молекул CD4 или CD8 утрачивается и клетки становятся зрелыми. Клетки сохранившие CD4 являются Т-хелперами (Т h) и их ТКР распознает HLA II класса, а сохранившие CD8 – цитотоксическими Т-лимфоцитами и их ТКР обладает способностью распознавать HLA I класса. Из тимуса они мигрируют в периферические лимфоидные органы, где заселяют преимущественно Т-зависимые зоны. В частности в лимфоузлах – паракортикальную. Зрелые лимфоциты рециркулируют.
Таким образом, АНТИГЕННЕЗАВИСИМАЯ дифференцировка Т‑лимфоцитов включает в себя пролиферацию, приобретение специфических маркеров Т‑лимфоцитами и образование дифференцированных, зрелых субпопуляций, способных осуществлять характерные для той или иной субпопуляции функции (индукция иммунного ответа, его регуляция, цитотоксичность). В процессе антигеннезависимой дифференцировки образуются лимфоциты, которые генетически детерминированы к взаимодействию с определенным антигеном и иммунному ответу на этот антиген.
АНТИГЕНЗАВИСИМАЯ ДИФФЕРЕНЦИРОВКА Т-ЛИМФОЦИТОВ
Антигензависмая дифференцировка происходит в периферических органах иммунной системы, если Т-лимфоцит провзаимодействовал с антигеном. Начинается она с момента распознавания антигена и заканчивается образованием клона лимфоцитов, способных оказывать специфическое действие как по отношению к антигену, так и к другим иммунокомпетентным клеткам, вступившим во взаимодействие с антигеном. Причем хелперы и цитотоксические лимфоциты распознают антиген по-разному. Так, ХЕЛПЕРЫ (CD4–клетки) распознают АНТИГЕН в комплексе с HLA II КЛАССА, КИЛЛЕРЫ (CD8-клетки) - в комплексе антиген с HLA 1 КЛАССА. Распознавание антигена Т‑хелпером является центральным процессом, как в гуморальном иммунном ответе, так и в усилении клеточной формы иммунного ответа.
Специфическими МАРКЕРАМИ для ВСЕЙ ПОПУЛЯЦИИ Т‑ЛИМФОЦИТОВ являются имеющиеся на наружной мембране этих клеток антигены CD 3. (Раньше использовался маркер CD 2 - рецептор для эритроцитов барана, что не совсем корректно. Параметры CD антигенов смотри в приложении.)
Маркер Т-лимфоцитов - структура, которая свойственна только Т‑лимфоцитам (всем субпопуляциям Т-лимфоцитов) – CD3.
|
| ||||||
| |||||||
| |||||||
| |||||||
Субпопуляции лимфоцитов:
Т h лимфоциты. Примерно половина из числа циркулирующих Т‑лимфоцитов несут на своей поверхности антиген CD4 . Эти Т‑лимфоциты функционируют как ХЕЛПЕРЫ, то есть помощники (от англ. to help — помогать), «включающие» популяцию В‑лимфоцитов в процесс выработки антител, а Т-эффекторы – в реализацию клеточного иммунитета. Т‑хелперы опосредуют свою функцию гуморальными факторами - цитокинами, которые синтезируются этими лимфоцитами в ответ на антигенный стимул.
Недостаточность хелперной функции Т‑лимфоцитов, наблюдаемая при синдроме приобретенного иммунодефицита (СПИД, одной из важнейших мишеней ВИЧ являются Т-лимфоциты хелперы), приводит к «неотвечаемости» организма на антигенную стимуляцию, что, в конечном итоге, способствует персистенции в организме человека микроорганизмов, развитию злокачественных новообразований и является причиной летального исхода.
Т-хелперы ( Th) – стимулируют пролиферацию и дифференцировку как Т-, так и В-лимфоцитов, выделяя цитокины. В зависимости от того, какие цитокины они продуцируют (в зависимости от цитокинового профиля) среди них различают:
Th1 (Т-хелперы первого типа), выделяют ИЛ-2 и γ-интерферон, и в итоге обеспечивают реакции Т-клеточного иммунитета – стимулируют иммунный ответ против внутриклеточных бактерий, противовирусный, противоопухолевый, трансплантационный иммунитет.
Th2 (Т-хелперы второго типа), секретируют ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10, ИЛ-13 и стимулируют синтез антител, способствуют развитию гуморального иммунного ответа против внеклеточных бактерий, их токсинов, а также образование IgE-антител.
Между Th1 и Th2 существует антагонизм: при повышении активности одних, угнетается функция других. В итоге преобладает Т-клеточный (Th1Ø T киллеры) или В-клеточный (Th2 Ø В-лимфоциты Ø антитела) иммунитет, что во многом зависит от вида антигена. Таким образом, Т‑хелперы выполняют хелперно-регуляторную функцию во взаимодействии иммунокомпетентных клеток, направленную на развитие эффекторной фазы иммунного ответа. Именно от Th зависит будет преобладать гуморальный или клеточный иммунный ответ.
|
Т k. Среди субпопуляций Т‑лимфоцитов различают эффекторные клетки. В связи с тем, что эти эффекторные клетки способны специфически разрушать клетки-мишени, их называют ЦИТОТОКСИЧЕСКИМИ Т‑ЛИМФОЦИТАМИ, или Т‑КИЛЛЕРАМИ - убийцами (от англ. to kill - убивать). Т‑киллер —одна из основных эффекторных клеток клеточноопосредованного иммунитета, которая наряду с другими клетками способна осуществлять лизис клеток мишеней. Роль Т‑киллеров очень велика в реализации трансплантационного иммунитета, развитии аутоиммунных заболеваний, в противоопухолевой защите. Тk-лимфоциты (CD8+ клетки) составляют около 20 – 25% от числа циркулирующих Т-лимфоцитов (абсолютное количество - 500 – 1200 в 1 мм3 (мкл)), они несут маркерный антиген CD8 . Макромолекула CD8 служит корецептором для антигенов главного комплекса гистосовместимости I класса (ГКГС-1).
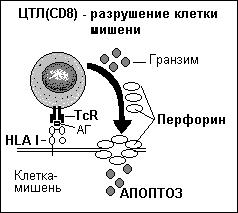 Активированные антигеном цитотоксические клетки – Т-киллеры связываются с антигенами на поверхности клеток и, выделяя белок перфорин, разрушают их. При этом Т-киллер остается жизнеспособным и может разрушать следующую клетку. Действие перфорина подобно МАК системы комплемента. Белок перфорин, полимеризуясь в мембране клетки-мишени, образует поры – каналы, тем самым, вызывая её осмотический лизис. Кроме того, цитотоксический Т-лимфоцит через пору, образованную перфорином в клетке-мишени, вбрасывает гранзимы (ферменты – сериновые протеазы), которые запускают программу апоптоза. Установлено также, что своё цитотоксическое действие Т-лимфоциты могут реализовывать путем экспрессии FasL и с его помощью индуцировать Fas-опосредованный апоптоз мишени.
Активированные антигеном цитотоксические клетки – Т-киллеры связываются с антигенами на поверхности клеток и, выделяя белок перфорин, разрушают их. При этом Т-киллер остается жизнеспособным и может разрушать следующую клетку. Действие перфорина подобно МАК системы комплемента. Белок перфорин, полимеризуясь в мембране клетки-мишени, образует поры – каналы, тем самым, вызывая её осмотический лизис. Кроме того, цитотоксический Т-лимфоцит через пору, образованную перфорином в клетке-мишени, вбрасывает гранзимы (ферменты – сериновые протеазы), которые запускают программу апоптоза. Установлено также, что своё цитотоксическое действие Т-лимфоциты могут реализовывать путем экспрессии FasL и с его помощью индуцировать Fas-опосредованный апоптоз мишени.
«Наивные» Т-лимфоциты – это те лимфоциты, которые не встретились с антигеном и они составляют часть общего пула рециркулирующих Т-клеток.
Т-клетки иммунологической памяти – это долгоживущие лимфоциты, потомки клеток, встречавшихся с антигенами и сохраняющие к ним рецепторы. Т‑ЛИМФОЦИТЫ ИММУНОЛОГИЧЕСКОЙ ПАМЯТИ - после стимуляции антигеном способны сохранять информацию о нем до 10—15 лет и передавать ее другим клеткам. Эти клетки защищены от апоптоза. Благодаря наличию в организме Т‑клеток памяти обеспечивается ускоренный иммунный ответ по вторичному типу при повторном попадании данного антигена в организм. Этим объясняется форсированная динамика вторичного иммунного ответа. Маркером Т-лимфоцитов памяти является мембранный антиген CD45RO.
Раньше ошибочно выделяли субпопуляцию Т-супрессоров, которые считались ответственными за подавление иммунного ответа. Однако в настоящее время показано, что самостоятельной субпопуляции Т-супрессоров нет. В угнетении, подавлении иммунного ответа решающее значение играет апоптоз простимулированных лимфоцитов, а также цитокин – трансформирующий фактор роста β.
Около 10 % лимфоцитов не имеют ни Т‑, ни В‑маркеров, они не относятся ни к Т‑, ни к В‑лимфоцитам и ранее получили название НУЛЕВЫЕ ЛИМФОЦИТЫ . Эта разнородная популяция лимфоцитов в зависимости от их морфофункциональных особенностей подразделяется на:
l ЕСТЕСТВЕННЫЕ (natural) КИЛЛЕРНЫЕ КЛЕТКИ (сокращенно ЕКК=ЕК=NK-клетки) и
l КИЛЛЕРНЫЕ КЛЕТКИ (К‑клетки).
Общей особенностью ЕК- и К‑клеток является способность лизировать клетки-мишени без предварительной сенсибилизации, что необходимо Т-лимфоцитам‑киллерам. Морфологически это лимфоциты большого размера с зернистой цитоплазмой. Дифференцируются из общей клетки предшественника лимфоцитов (LSC).
Естественные киллеры не зависят в своем развитии от вилочковой железы. Экспрессируют на своей поверхности рецепторы к интерферону-γ и интерлейкину‑2 (ИЛ 2). Функционально они являются цитотоксическими клетками киллерами, но на NK нет антигенраспознающих рецепторов, которые обязательно присутствуют на Т-киллерах. Натуральных киллеров на клетку мишень наводят антитела IgG специфичные к мембранным антигенам клетки-мишени. Первоначально антитела связываются с антигеном на клетке, а затем с помощью Fc рецептора к IgG (FcγRIII) NK присоединяется к этому комплексу АТ - АГ-клетки-мишени. Функция ЕК-клеток в организме заключается в защите от развития опухолей, вирусов и др. Основными их маркерами являются CD16 и CD56. (FcγRIII по CD- номенклатуре это CD16).Разрушение клетки-мишени ЕК осуществляет с помощью перфорина. Содержание ЕК (CD16+ клеток) у здоровых людей – 8 – 22%.
 К‑клетки неоднородная группа клеток, несущая на своей поверхности рецепторы к Fc- фрагменту Ig G и способны к антителозависимой клеточной цитотоксичности. К ним относятся моноциты, нейтрофилы, макрофаги, эозинофилы, конечно НК и некоторые лимфоциты. Антителозависимая клеточноопосредованная цитотоксичность (АЗКЦ) является своеобразным отражением связи между гуморальным и клеточным звеньями иммунной системы. Антитела выступают в роли «наводчиков» клеток-эффекторов на клетки-мишени, несущие чужеродные антигены.
К‑клетки неоднородная группа клеток, несущая на своей поверхности рецепторы к Fc- фрагменту Ig G и способны к антителозависимой клеточной цитотоксичности. К ним относятся моноциты, нейтрофилы, макрофаги, эозинофилы, конечно НК и некоторые лимфоциты. Антителозависимая клеточноопосредованная цитотоксичность (АЗКЦ) является своеобразным отражением связи между гуморальным и клеточным звеньями иммунной системы. Антитела выступают в роли «наводчиков» клеток-эффекторов на клетки-мишени, несущие чужеродные антигены.
Все лимфоциты (Т‑, В‑, ЕК- и К‑клетки) обладают способностью к миграции и рециркуляции (см. методическую рекомендацию к первому занятию), что обеспечивает повсеместный контроль за размножением клеток собственного организма, а при проникновении чужеродного антигена —генерализированный иммунный ответ и сохранение иммунологической памяти об антигене.
МАРКЕРЫ ЛИМФОЦИТОВ
 |
ПРОТОЧНАЯ ЦИТОФЛЮОРОМЕТРИЯ позволяет определить с помощью моноклональных антител, меченных флюорохромом, маркерные CD-антигены на поверхности клеток и таким образом выявлять популяции и субпопуляции лимфоцитов, степень их активности и функциональной активности.
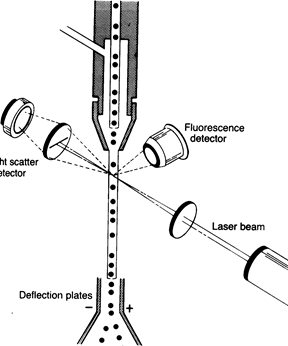
 Принцип проточной цитофлюорометрии. Меченная флюоресцентными моноклональными антителами исследуемая клетка проходит в потоке жидкости в капилляре. Поток жидкости пересекается лучом лазера и прибор фиксирует отраженный от поверхности клетки сигнал по принципу да/нет (есть клетка или нет). Наличие на клетке связанных с её дифференцировочными антигенами моноклональных антител, меченных флюорохромом, указывает о принадлежности клетки к определенной субпопуляции.
Принцип проточной цитофлюорометрии. Меченная флюоресцентными моноклональными антителами исследуемая клетка проходит в потоке жидкости в капилляре. Поток жидкости пересекается лучом лазера и прибор фиксирует отраженный от поверхности клетки сигнал по принципу да/нет (есть клетка или нет). Наличие на клетке связанных с её дифференцировочными антигенами моноклональных антител, меченных флюорохромом, указывает о принадлежности клетки к определенной субпопуляции.
Определение CD3-клеток (Т-лимфоцитов), CD19 (В-лимфоцитов) в крови имеет диагностическое значение при первичных и вторичных иммунодефицитах. Важную роль CD-типирование Т-клеток играет при лимфопролиферативных за-болеваниях (лимфо-лейкозах), реакциях отторжения трансплантата и РТПХ (реакции трансплантат против хозяина), вирусных и бактериальных инфекциях.
Диагностическую значимость имеет определение CD4-лимфоцитов в крови при иммунодефицитах, как гуморального, так и клеточно-опосредованного иммунитета. Необходимо подчеркнуть, что количество CD4-клеток является решающим показателем для прогноза развития СПИД у ВИЧ инфицированных. Определение индекса CD4/CD8 (соотношения количества хелперов к эффекторам), так называемого индекса регуляции, важно при ВИЧ-инфекции. Так, снижение CD4 до 500/мкл и ниже считается клиническим стандартом для начала антиретровирусной терапии, а снижение их количества до 200/мкл и ниже — для начала профилактической терапии оппортунистических инфекций.
Приложение.
 |
(Приводим параметры CD антигенов. Это справочная информация, которая поможет Вам при чтении литературы по иммунологии, иммунопатологии, гематологии. Значимые CD антигены отмечены v. С ними Вы встречались на предыдущих занятиях, будут разбираться на текущем и будущих.)
· CD1 - a, b, c; его несут кортикальные тимоциты, субпопуляции В-клеток, клетки Лангерганса, является общим антигеном тимоцитов, белок, подобен антигенам 1класса гистосовместимости, ММ 49 КД.
· v CD2 - маркер всех Т-клеток, имеют также большинство (~ 75%) ЕК, известны три эпитопа молекулы, один из которых связывает эритроциты барана (Е-рецептор); является адгезивной молекулой связывается с CD58 (LFA III), LFA IV, передает трансмембранные сигналы при активации Т-клеток; ММ 50 КД. Выявить этот антиген можно реакцией розеткообразования. Реакция Е-розеткообразования является показателем суммы клеток (Т-л, ЕК, LAK), несущих CD2-кластер. Таким образом, CD2 –антиген не является абсолютным маркером Т-лимфоцитов, так как присутствует и на других клетках.
· v CD3 – несут все зрелые Т-лимфоциты, обеспечивает передачу сигнала от Т-клеточного антигенспецифического рецептора (ТКР) в цитоплазму, состоит из пяти полипептидных цепей (γ, δ, ε, ι, ξ). ММ – 25 КД; антитела к нему усиливают или ингибируют функцию Т-клеток. Важный маркёр Т-лимфоцитов.
· v CD4 – маркер Т-хелперов, рецептор к вирусу иммунодефицита человека (ВИЧ), имеется на некоторых моноцитах, клетках глии; трансмембранный гликопротеин, участвует в распознавании антигенов, ассоциированных с молекулами II класса гистосовместимости (HLA-DR), ММ 59 КД. (Рецептор для АГ ГКГС класса ll).
· v CD5 – имеют зрелые и незрелые Т-клетки, трансмембранный гликопротеин, член семейства рецепторов – “мусорщиков”, как и CD6, является лигандом для CD72 на В-клетках, участвует в пролиферации Т-клеток. CD5 так же имеют В-1-лимфоциты – субпопуляция В-клеток, с преимущественной локализацией в брюшной и плевральной полостях. ММ 67 КД.
· CD6 – несут зрелые Т-клетки и частично В-клетки имеют все Т-клетки и тимоциты, часть В-клеток; входит в семейство «мусорщиков», ММ 120 КД.
· CD7 – имеют Т-клетки, ЕК (Fcμ рецептор IgM); ММ 40 КД.
· v CD8 – маркер цитотоксических Т-лимфоцитов, имеют некоторые ЕК, структура адгезии, вовлекается в распознавание антигенов при участии молекул гистосовместимости 1 класса, состоит из двух S-S цепей , ММ 32 КД. (Корецептор для комплекса АГ + ГКГС класса l).
· CD9 – несут моноциты, тромбоциты, гранулоциты, В-клетки фолликулярных центров, эозинофилы, базофилы, эндотелий, ММ 24 КД.
· CD10- имеют незрелые В-клетки (GALLA – антиген лейкозных клеток), часть тимоцитов, гранулоцитов; эндопептидаза, ММ 100 КД.
· CD11 a – несут все лейкоциты, молекула цитоадгезии, αL цепь интегрина LFA-1, ассоциирована с CD18; рецептор для лигандов: CD15 (ICAM-1), CD102 (ICAM-2) и CD50 (ICAM-3) молекул; отсутствует у больных с LAD-1 синдромом (синдром дефицита молекулы адгезии), ММ 180 КД.
· v CD11 b – (CR3- или c3bi-рецептор) – несут моноциты, гранулоциты, ЕК, αМ цепь интегрина, ассоциирована с CD18молекулой; рецептор для лигандов: CD54 (ICAM-1), C3bi-компонента комплемента (CR3-рецептор) и фибриногена; отсутствует при LAD-1 синдроме: ММ 165 КД.
· v CD11 c (CR4-рецептор) – имеют моноциты, гранулоциты, ЕК, активированные Т- и В-лимфоциты, αХ цепь интегрина (ассоциирована с CD18, является четвертым типом рецептора (CR4) для компонентов C3bi, C3dg комплемента; его лиганды– СD54 (ICAM-1), фибриноген; ММ 95/150 кД.
· CD13 – имеют все миелоидные, дендритные и эндотелиальные клетки, аминопептидаза N, рецептор для коронавируса, ММ 150 КД.
· v CD14 – имеют моноциты/макрофаги, гранулоциты, рецептор для комплексов ЛПС с ЛПС-связывающим белком и для PI-молекул тромбоцитов; отсутствует у больных с пароксизмальной ночной гемоглобинурией (PNH), антитела к нему могут вызывать окислительный взрыв в моноцитах, ММ 55 КД.
· CD15 – (Lewis) – имеют гранулоциты, слабо экспрессируют моноциты, некоторые антитела к нему подавляют фагоцитоз.
· CD15 s – (sialyl-Lewis) – имеют миелоидные клетки, лиганд для CD62P (P-селектин), CD62E (E-селектин), CD62L (L-селектин), отсутствует у больных LAD-2.
· v CD16 – несут ЕК, нейтрофилы, некоторые моноциты, (низкоаффинный Fc-рецептор для IgG), интегральный мембранный белок (Fcγ RIIIA) на ЕК и макрофагах, PI-связывающая форма (Fcγ RIIIB) на нейтрофилах, отсутствует у больных с PNH – пароксизмальной ночной гемоглобинурией.
· CD18 – имеют большинство лимфоидных и миелоидных клеток, молекула адгезии, β2 цепь интегрина LFA, ассоциирован с αCD11 a, b, c, отсутствует при LAD-1 синдроме, ММ 95 КД.
· v CD19 – (В4) – имеют пре-В и В-клетки, часть их рецепторного комплекса вовлекается в их активацию (сигнал трансдукции, ассоциирован с CD21 (CR2); ММ 95 КД. Важный маркёр В-клеток.
· v CD20 – (В1) – несут все В-клетки и дендритные клетки в фолликулах, участвует в активации через кальциевые каналы клеток, ММ 35 кДа.
· v CD21 – (CR2 рецептор, В2) - имеют субпопуляции В-клеток, некоторые тимоциты, Т-клетки, рецептор для С3d компонента комплемента и для вируса Эпштейна-Барра, участвует в регуляции активации комплемента (RCA) наряду с CD35, CD46 CD55 и в активации В-клеток.
Более полную информацию о кластерах дифференцировки можно найти в учебниках 1 и 2 списка литературы для самоподготовки на с.16.
Показатели содержания лимфоцитов у здоровых людей
| Популяции лимфоцитов и маркеры | Т-имфоци-ты | Т-хелперы | Т-цитоток-сические | В-лимфо-циты | Естествен-ные киллеры |
| CD3 | CD4 | CD8 | CD19 | CD16 | |
| Процентное содержание (%) | 60-80 | 30-50 | 20-25 | 10-23 | 8-22 |
| Абсолютное количество в 1 мкл | 800-2200 | 500-1200 | 300-600 | 150-600 | 100-600 |
Индекс регуляции CD4/CD8 - 1,2-2,5. * мкл = 1 мм3.
Дата добавления: 2021-07-19; просмотров: 68; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!