Механизм перемещения веществ по эндоцитозному пути остается недостаточно понятным и описывается двумя моделями.
Разновидности ЭПС
-Гранулярная ЭПС
-Агранулярная (гладкая) ЭПС
-Переходная (транзиторная) ЭПС
4. Гранулярная (шероховатая) эндоплазматическая сеть. ЭМ, функции. Особенности трансляции на грЭПС (фолдинг, специфическая сортировка, модификация и транспорт белков).
Гранулярная ЭПС обеспечивает:
(1) биосинтез всех мембранных белков и белков, предназначенных для экспорта из клетки, и
(2) начальное гликозилирование и посттрансляционные изменения белковых молекул.
Гранулярная ЭПС образована уплощенными мембранными цистернами и трубочками, на наружной поверхности которых располагаются рибосомы и полисомы, придающие мембранам зернистый (гранулярный) вид (см. рис. 3-7 и 3-8), что и отражено в названии органеллы.
Мембраны грЭПС содержат особые белки, которые обеспечивают
(1) связывание рибосом и (
2) уплощение цистерн.
Полость грЭПС содержит рыхлый материал умеренной плотности (продукты синтеза) и сообщается с перинуклеарным пространством (см. ниже).
Благодаря грЭПС происходит отделение (сегрегация) вновь синтезированных белковых молекул от шалоплазмы.
СИНТЕЗ БЕЛКА НА ГРЭПС начинается на свободных полисомах, которые в дальнейшем связываются с мембранами ЭПС (см. рис. 3-8).
На первом этапе взаимодейстия иРНК с рибосомами происходит образование особого сигнального пептида (длиной 20-25 аминокислот), связывающегося с рибонуклеопротеидным комплексом - сигнал-распознающею частицей (СРЧ). Присоединение СРЧ к сигнальному пептиду угнетает дальнейший синтез белка до тех пор, пока комплекс СРЧ-полисома не свяжется со специфическим рецептором на мембране ЭПС - причальным белком (docking protein в англоязычной литературе).
|
|
|
После связывания с рецептором СРЧ отделяется от полисом, что разблокирует синтез белковой молекулы. В мембране грЭПС имеются интегральные рецепторные белки рибофорины, обеспечивающие прикрепление больших субъединиц рибосом.
Эти белки не диффундируют в область аЭПС и формируют гидрофобные каналы в мембране, служащие для проникновения вновь синтезированной белковой цепочки в просвет грЭПС, что, наряду с рибофоринами, способствует удержанию рибосом на поверхности мембран грЭПС.
В просвете грЭПС сигнальный пептид отщепляется особым ферментом сигнальной пептидазой, которая располагается на внутренней поверхности мембраны. В ходе продолжающейся трансляции внутри цистерны грЭПС накапливается белок, который приобретает вторичную и третичную структуру, а также подвергается начальным посттрансляционным изменениям - гидроксилированию, сульфатированию и фосфорилированию.
|
|
|
Наиболее важным из этих изменений является гликозилирование - присоединение к белкам олигосахаридов с образованием гликопротеинов, которое происходит перед секрецией или транспортом большинства белков к другим участкам внутри клетки (комплексу Гольджи, лизосомам или плазмолемме). В отличие от них, растворимые белки гаалоплазмы не гликозилированы. Гликозилирование обеспечивается связанным с мембраной ферментом гликозилтрансферазой, переносящим олигосахарид.
Хотя грЭПС присутствует во всех клетках (за исключением спермиев), степень ее развития существенно варьирует. Она особенно хорошо развита в клетках, специализирующихся на белковом синтезе, например, в эпителиальных железистых клетках ацинусов поджелудочной железы (вырабатывающих пищеварительные ферменты), фибробластах (синтезирующих коллаген и ряд других белков), плазматических клетках (продуцирующих иммуноглобулины). Для вех этих клеток характерна выраженная базофилия цитоплазмы в области расположения элементов грЭПС. В нейронах отдельным компактным скоплениям цистерн грЭПС на светооптическом уровне соответствуют очерченные участки базофилии цитоплазмы, которые в совокупности называются хроматофильной субстанцией или тельцами Ниссля.
|
|
|
5. Агранулярная (гладкая) эндоплазматическая сеть. ЭМ, функции .
Агранулярная (гладкая) ЭПС представляет собой трехмерную замкнутую сеть мембранных анастомозирующих трубочек, канальцев, цистерн и пузырьков диаметром 20-100 нм, на поверхности которых рибосомы отсутствуют (см. рис. 3-7), что определило ее название.
Соответственно, на мембранах аЭПС отсутствуют рецепторы, связывающие субъединицы рибосом (рибофорины).
Предполагают, что аЭПС образуется в результате формирования выростов грЭПС, мембрана которых утрачивает рибосомы.
Функции аЭПС включают:
(I) синтез липидов, в том числе мембранных (ферменты липидною синтеза располагаются на наружной - обращенной в сторону гаалоплазмы - поверхности мембраны аЭПС),
(2) синтез гликогена,
(3) синтез холестерина,
(4) детоксикацию эндогенных и экзогенных веществ,
(5) накопление ионов Са2 + ,
(6) восстановление кариолеммы в телофазе митоза (эта функция оспаривается авторами, считающими, что кариолемма восстанавливается за счет мембранных пузырьков, на которые она ранее распалась).
Помимо указанных основных функций, в некоторых типах клеток аЭПС выполняет ряд дополнительных - например, в мегакариоцитах (гигантских клетках костного мозга) ее элементы образуют демаркационные каналы, разделяющие формирующиеся тромбоциты.
|
|
|
Способность аЭПС к накоплению ионов Са2 + обусловлена наличием:
(1) кальциевого насоса в ее мембране, который обеспечивает транспорт этих ионов из гиалоплазмы внутрь цистерн аЭПС;
(2) кальций-связывающих белков (кальсеквестрина в мышечных клетках, кальретикулина - преимущественно в немьшечных и др.), которые в просвете цистерн образуют комплекс с ионами Са2 + и
(3) кальциевых каналов в мембране аЭПС, которые осуществляют выведение Са2 + в гиалоплазму.
Механизмы действия кальциевых каналов неодинаковы в клетках разных типов. Функция накопления ионов Са2 + особенно выражена в мышечных клетках, в которых специализированная аЭПС (именуемая саркоплазматической сетью) обеспечивает мышечное сокращение путем накопления и выделения значительных количеств ионов Са2+, связывающихся с особыми белками.
Обычно аЭПС в цитоплазме занимает меньший объем, чем грЭПС, однако она очень хорошо развита в клетках, синтезирующих стероиды, триглицериды и холестерин. Так, аЭПС занимает значительную часть объема цитоплазмы в клетках, которые активно продуцируют стероидные гормоны (клетки коркового вещества надпочечника, интерстициальные гландулоциты яичка (клетки Лейдига), клетки желтого тела яичника (лютеоциты) и др.
Она также хорошо развита в клетках печени (гепатоцитах), где ее ферменты участвуют в процессах окисления, конъюгации и метилирования, которые обеспечивают нейтрализацию и детоксикацию ряда гормонов и вредных веществ (алкоголя, инсектицидов и др.).
6. Комплекс Гольджи (пластинчатый аппарат). Структурная организация при СМ и ЭМ. Полярность комплекса Гольджи. Функции, участие в процессинге молекул и транспорте веществ.
Комплекс Гольджи - сложно организованная мембранная органелла, образованная тремя основными элементами –
(1) стопкой уплощенных мешочков (цистерн),
(2) пузырьками и
(3) вакуолями, или секреторными пузырьками (см. рис. 3-1 и 3-9).
Комплекс этих элементов называется диктиосомой (от греч. с11к1уоп - сеть); в некоторых клетках имеются множественные диктиосомы (до нескольких сотен). В специализированных секреторных клетках комплекс Гольджи располагается надъядерно под апикальной частью клетки, через которую происходит выделение секрета механизмом экзоцитоза. Нередко он лежит у ядра вблизи центриолей, в некоторых клетках его компоненты рассеяны по всей цитоплазме.
1. Цистерны Каждая группа цистерн внутри стопки отличается особым составом ферментов, определяющим характер реакций процессинга белков. Периферические отделы цистерн несколько расширены, от них отщепляются пузырьки и вакуоли. Механизм, удерживающий стопку в виде единого образования, неизвестен. При наличии в клетке множественных диктиосом их цистерны связаны друг с другом системой анастомозирующих и ветвящихся трубочек.
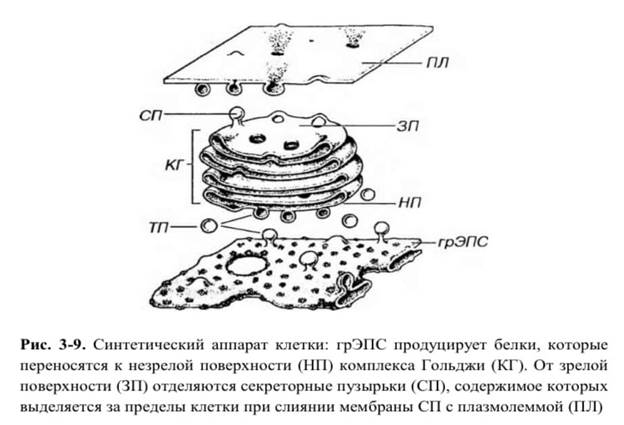
2. Пузырьки - сферические окруженные мембраной элементы диаметром 40-80 нм с содержимым умеренной плотности; образуются путем отщепления от цистерн
3. Вакуоли - крупные (диаметр - 0.1-1.0 мкм), окруженные мембраной сферические образования, отделяющиеся от цистерны на зрелой поверхности комплекса Гольджи (см. ниже) в некоторых железистых клетках. Они содержат секреторный продукт умеренной плотности, находящийся в процессе конденсации (конденсирующие вакуоли).
Полярность комплекса Гольджи. Комплекс Гольджи представляет собой поляризованную структуру, в которой выделяют две поверхности, обладающие структурными и функциональными различиями: (а) цис- (от лат. а я - по эту сторону), незрелую, формирующуюся - выпуклой формы, обращенную к ЭПС и связанную с системой мелких (транспортных) пузырьков, отщепляющихся от ЭПС; - 51 - (б) транс- (от лат. trans - по ту сторону), зрелую - вогнутой формы, обращенную к плазмолемме и связанную с отделяющимися от цистерн вакуолями. Между цистернами цис- и транс-поверхностей располагаются цистерны медиальной части комплекса Гольджи.
Транспорт веществ в комплексе Гольджи. Белки проникают в стопку цистерн комплекса Гольджи из транспортных пузырьков с цис- поверхности, а выходят в вакуолях с транс-поверхности; каким образом осуществляется их перенос внутри комплекса, в ходе которого происходит их процессинг, остается неизвестным.
Возможные пути этого транспорта описываются двумя моделями:
1) модель перемещения цистерн постулирует, что за счет слияния транспортных пузырьков на цис-поверхности непрерывно происходит новообразование цистерн (что легло в основу термина "формирующаяся поверхность"), в дальнейшем смещающихся к транс-поверхности, по достижении которой они распадаются на вакуоли ("зрелая поверхность"). Согласно этой модели, одни операции процессинга сменяются другими при перемещении самой цистерны по ходу изменений ее состава. Транспорт веществ из одной цистерны в другую, в соответствии с описанной моделью, отсутствует;
2) модель везикулярного транспорта предполагает, что цистерны не меняют своего расположения (остаются постоянно на своем месте), а продукты синтеза переносятся от цис- к транс-поверхности в пузырьках (везикулах), которые отпочковываются от предшествующей цистерны, сливаясь с последующей.
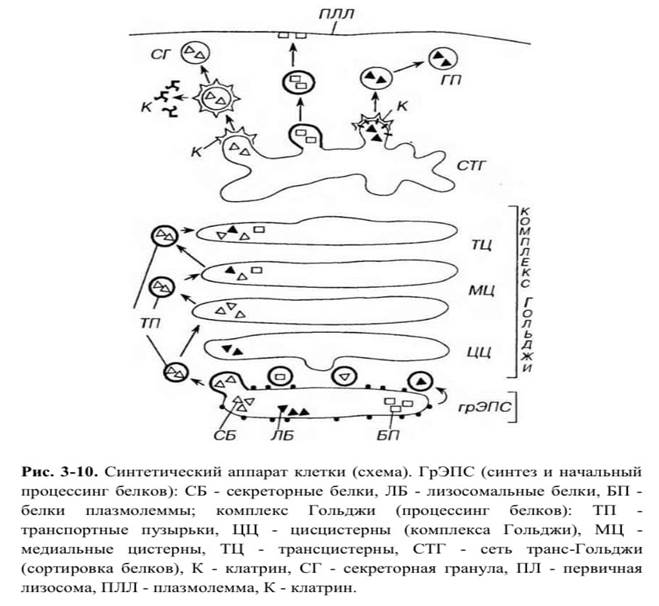
Функции комплекса Гольджи: 1) синтез полисахаридов и гликопротеинов (гликокаликса, слизи); 2) процессинг молекул: включение углеводных компонентов в гликопротеины, транспортируемые из грЭПС (терминальное гликозилирование), добавление фосфатных групп (фосфорилирование), жирных кислот (ацилирование), сульфатных остатков (сульфатирование), частичное расщепление белковых молекул (протеолитическая доработка). Каждый их указанных этапов процессинга веществ внутри комплекса Гольджи осуществляется в топографически определенном его компоненте (цис-, медиальных или транс-цистернах, а также сети транс-Голь- джи); 3) конденсация секреторного продукта (в конденсирующих вакуолях) и образование секреторных гранул; 4) обеспечение новообразованных гранул мембраной (синтезированной в ЭПС) и упаковка в нее секреторных продуктов; в процессе секреции эта мембрана встраивается в плазмолемму, увеличивая площадь ее поверхности; - 52 - 5) сортировка белков на транс-поверхности (в сети транс-Голъджи) перед их окончательным транспортом. Направление последующего транспорта различных белков из комплекса Гольджи зависит от особенностей их гликозшшрования, фосфорилирования и сульфатирования. Сортировка производится посредством специфических мембранных рецепторных белков, которые распознают сигнальные участки на макромолекулах и направляют их в соответствующие пузырьки.
Транспорт белков из комплекса Гольджи осуществляется в составе трех важнейших потоков (рис. 3-10): (1) в гидролазные пузырьки (ранее называемые первичными лизосомами) - начально в виде окаймленных пузырьков, (2) в плазмолемму (в составе окаймленных пузырьков) и (3) в секреторные гранулы (в виде окаймленных пузырьков, утрачивающих в дальнейшем оболочку).
7. Структурно-химическая организация эндосом и лизосом (СМ и ЭМ). Функции. Типы лизосом.
АППАРАТ ВНУТРИКЛЕТОЧНОГО ПЕРЕВАРИВАНИЯ представлен системой особых органелл - мембранных пузырьков с кислым содержимым - эндосом (от греч. endo - внутри и soma - тело) и лизосом (от греч. lysis - разрушение и soma - тело), которые обеспечивают катаболические процессы в цитоплазме клетки (рис. 3-11).
Функция аппарата внутриклеточного переваривания состоит в регулируемом внутриклеточном расщеплении макромолекул внеклеточного и внутриклеточного происхождения.
Эндосомы - мембранные пузырьки с постепенно закисляющимся содержимым, которые обеспечивают перенос макромолекул с поверхности клетки в лизосомы и их частичный или полный гидролиз на стадиях, предшествующих лизосомальному уровню деградации.
В связи с указанными свойствами совокупность эндосом в настоящее время считают не просто механизмом транспорта веществ в клетке (как полагали ранее), а частью системы их переваривания ("внутриклеточного пищеварительного тракта"), в которую входят также лизосомы.
Процесс переноса веществ системой эндосом (по эндоцитозному пути) может протекать
(а) с полным перевариванием макромолекул,
(б) с их частичным расщеплением или
(в) без изменений по ходу транспорта в лизосому.
Способность к перевариванию в эндосомах обеспечивается благодаря тому, что кислые гидролазы вносятся в эндоцитозный путь уже на самых ранних его этапах.
Путь транспорта и деградации веществ в клетке можно описать последовательностью;
ранняя (периферическая) эндосома -> поздняя (перинуклеарная) эндосома -> лизосома.
Условия расщепления макромолекул на указанном пути их переноса последовательно становятся все более жесткими.
Эндосомы обеспечивают сравнительно мягкий контролируемый прелизосомальный этап переваривания, который необходим и достаточен для легко расщепляемых веществ и комплексов. Наибольшая активность и степень деградации веществ характерна для лизосом, куда переносятся наименее перевариваемые материалы. Благодаря такому устройству клетка располагает органеллами с широким спектром условий расщепления веществ.
Механизм перемещения веществ по эндоцитозному пути остается недостаточно понятным и описывается двумя моделями.
(1) модель челночных пузырьков основана на представлении о переносе поглощенных веществ между стабильными органеллами посредством транспортных пузырьков;
(2) модель созревания предполагает последовательное превращение ("созревание") одной органеллы в другую в пределах указанного пути (компоненты, необходимые для процесса созревания, например, гидролазы, доставляются пузырьками, сливающимися с созревающими эндо-сомами).
Ранние (периферические) эндосомы являются мембранными пузырьками на ранних этапах после их отделения от плазмолеммы (но уже после утраты первоначально имевшейся клатриновой оболочки). Они располагаются неподалеку от плазмолеммы в периферических отделах цитоплазмы (см. рис. 3-11). В них в условиях слабокислой среды (pH 6.0) осуществляется ограниченное и регулируемое переваривание макромолекул протеазами, которые были внесены в эндосому, по-видимому, еще на этапе ее формирования
Поздние (перинуклеарные) эндосомы получили свое название благодаря тому, что они образуются позднее ранних и располагаются в глубоких отделах цитоплазмы вблизи ядра. Они достигают диаметра 600-800 нм и характеризуются сравнительно плотным матриксом. Их отличает от ранних эндосом более кислое содержимое (pH 5.5) и более глубокий уровень переваривания ферментами. В них из ранних эндосом поступают продукты (лиганды), которые должны подвергнуться расщеплению. Большая часть этих продуктов, а также ферменты в дальнейшем будут направлены в лизосому (см. рис. 3-11). Предполагают, однако, что некоторые молекулы могут рециклироваться из поздней эндосомы в раннюю или даже направляться в плазмолемму.
Лизосомы (ранее называемые вторичными лизосомами) - органеллы, активно участвующие в завершающих этапах процесса внутриклеточного переваривания захваченных клеткой макромолекул посредством широкого спектра литических ферментов при низких значениях pH (5.0 и ниже). Они формируются с участием поздних эндосом. Диаметр лизосом обычно составляет 0.5-2 мкм, а их форма и структура могут существенно варьировать в зависимости от характера перевариваемого материала. Как и в случае гидролазных пузырьков, они достоверно идентифицируются только на основании выявления в них гидролитических ферментов.
Название отдельных видов лизосом основано на наличии в их просвете морфологически распознаваемого материала; в его отсутствие используется общий термин лизосома.
После переваривания содержимого лизосомы образующиеся низкомолекулярные вещества диффундируют через ее мембрану в гиалоплазму.
1) Фаголизосома формируется путем слияния поздней эндосомы или лизосомы с фагосомой, называемой также гетерофагосомой (от греч. heteros - другой, phagein - поедать и soma - тело) - мембранного пузырька, содержащего материал, захваченный клеткой извне и подлежащий внутриклеточному перевариванию; процесс разрушения этого материала называется гетерофагией;
2) Аутофаголизосома образуется при слиянии поздней эндосомы или лизосомы с аутофагосомой (от 1реч. autos - сам, phagein - поедать и soma - тело) - мембранным пузырьком, содержащим собственные компоненты клетки, подлежащие разрушению. Процесс переваривания этого материала называют аутофагией. Источником мембраны, окружающей клеточные компоненты, служит грЭПС.
3) Мулътивезикулярное тельце (от лат. multi - много и vesicula - пузырек) представляет собой крупную (диаметром 200-800 нм) сферическую окруженную мембраной вакуоль, содержащую мелкие (40-80 нм) пузырьки, погруженные с светлый или умеренно плотный матрикс. Оно образуются в результате слияния ранних эндосом с поздней, причем мелкие пузырьки формируются, вероятно, путем отпочковывания внутрь от мембраны вакуоли. Матрикс тельца содержит литические фермента и, очевидно, обеспечивает постепенное разрушение внутренних пузырьков.
4) Остаточные тельца - лизосомы, содержащие непереваренный материал, которые могут длительно находиться в цитоплазме или выделять свое содержимое за пределы клетки. Распространенным типом остаточных телец в организме человека являются липофусциновые гранулы - мембранные пузырьки диаметром 0.3-3 мкм, содержащие труднорастворимый коричневый эндогенный пигмент липофусцин. Под электронным микроскопом липофусциновые гранулы представляют собой структуры вариабельной формы, содержащие липидные капли, плотные гранулы и пластинки. В связи с их накоплением в некоторых клетках (нейронах, кардиомиоцитах) при старении, липофусцин рассматривают как "пигмент старения" или "изнашивания".
Секреция лизосомальных ферментов за пределы клетки осуществляется у остеокластов - клеток, разрушающих костную ткань, а также фагоцитов (нейтрофилов и макрофагов) при внеклеточном переваривании различных объектов. Избыточная секреция этих ферментов может приводить к повреждениям окружающих тканей.
8. Энергетическая система клетки. Митохондрии. Структурная организация (СМ и ЭМ). Митохондриальный матрикс. Функции митохондрий.
Митохондрии представляют собой мембранные полуавтономные органеллы, обеспечивающие клетку энергией, получаемой благодаря процессам окисления и запасаемой в виде фосфатных связей АТФ. Митохондрии также участвуют в биосинтезе стероидов, окислении жирных кислот и синтезе нуклеиновых кислот. Митохондрии могут иметь эллиптическую, сферическую, палочковидную, нитевидную и др. формы, которые могут изменяться в течение определенного времени. Их размеры составляют 0.2-2 мкм в ширину и 2-10 мкм в длину, а количество в различных клетках варьирует в широких пределах, достигая в наиболее активных 500-1000. В клетках печени (гепатоцитах) их число составляет около 800, а занимаемый ими объем равен примерно 20% объема цитоплазмы. На светооптическом уровне митохондрии выявляются в цитоплазме специальными методами и имеют вид мелких зерен и нитей (что обусловило их название - от греч. mitos- нить и chondros - зерно). В цитоплазме митохондрии могут располагаться диффузно, однако обычно они сосредоточены в участках максимального потребления энергии, например, вблизи ионных насосов, сократимых элементов (миофибрилл), органелл движения (аксонем спермия, ресничек), компонентов синтетического аппарата (цистерн ЭПС).
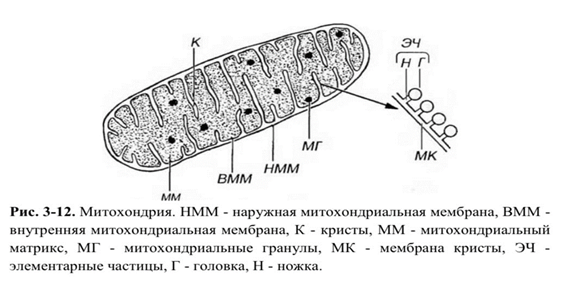
Митохондрии состоят из наружной и внутренней мембран, разделенных межмембранным пространством, и содержат митохондриальный матрикс, в который обращены складки внутренней мембраны - кристы (рис. 3-12)
Митохондриальный матрикс - гомогенное мелкозернистое вещество умеренной плотности, заполняющее полость (внутреннюю камеру) митохондрии и содержащее несколько сотен ферментов: растворимые ферменты цикла Кребса (за исключением СДГ, ферменты, участвующие в окислении жирных кислот, ферменты белкового синтеза. В матриксе находятся также митохондриальные рибосомы, митохондриальные гранулы и митохондриальная ДНК (что отличает митохондрии от всех остальных органелл).
Митохондриальные рибосомы имеют вид мелких плотных гранул, распределенных в матриксе. Белки, образующие эти рибосомы, лишь частично продуцируются в самой митохондрии. Митохондриальные гранулы - частицы высокой электронной плотности диаметром 20-50 нм с мелкозернистой или пластинчатой структурой, разбросанные по митохондриальному матриксу, содержащие ионы Са2 + и Mg2 + , а также другие дивалентные катионы. Функция гранул выяснена неполностью; предполагается, что их катионы необходимы для поддержания активности митохондриальных ферментов.
Митохондриальная ДНК (мтхДНК) - образует собственный геном митохондрий, на который приходится около 1% общего содержания ДНК в клетке и который включает 37 генов (в ядре клеток человека насчитывают примерно 100 тыс. генов). МтхДНК - кольцевой формы двунитчатая молекула ДНК длиной 5.5 мкм и толщиной 2 нм (в каждой митохондрии имеется 2-10 таких молекул). Она сходна с бактериальной ДНК и отличается от ядерной ДНК генетическим кодом, низким содержанием некодирующих последовательностей и отсутствием связи с гистонами.
9. Опорно-двигательная система клетки. Цитоскелет. Фибриллярные структуры цитоплазмы (микрофиламенты, промежуточные филаменты). Микротрубочки. ЭМ,
химический состав, функции.
Цитоскелет представляет собой сложную динамичную систему микротрубочек, микрофиламентов, промежуточных филаментов и микротрабекул. Указанные компоненты цитоскелета являются немембранными органеллами; каждый из них образует в клетке трехмерную сеть с характерным распределением, которая взаимодействует с сетями из других компонентов. Они входят также в состав ряда других более сложно организованных органелл (ресничек, жгутиков, микроворсинок, клеточного центра) и клеточных соединений (десмосом, полудесмосом, опоясывающих десмосом).
Основные функции цитоскелета: 1. поддержание и изменение формы клетки; 2. распределение и перемещение компонентов клетки; 3. транспорт веществ в клетку и из нее; 4. обеспечение подвижности клетки; 5. участие в межклеточных соединениях
Микрофиламенты - тонкие белковые нити диаметром 5-7 нм, лежащие в цитоплазме поодиночке, в виде сетей ими пучками. В скелетной мышце тонкие микрофиламенты образуют упорядоченные пучки, взаимодействуя с более толстыми миознновыми филаментами. Кортикальная (терминальная) сеть - зона сгущения микрофиламентов под плазмолеммой, характерная для большинства клеток. В этой сети микрофиламенты переплетены между собой и "сшиты" друг с другом с помощью особых белков, самым распространенным из которых является филамин. Кортикальная сеть препятствует резкой и внезапной деформации клетки при механических воздействиях и обеспечивает плавные изменения ее формы путем перестройки, которая облегчается актин-растворяющими (преобразующими) ферментами. - 70 - Прикрепление микрофиламентов к плазмолемме осуществляется благодаря их связи с её интегральными ("якорными") белками (интегринами) - непосредственно или через ряд промежуточных белков - талин, винкулин и αактинин (см. рис. 10-9). Помимо этого, актиновые микрофиламенты прикрепляются к трансмембранным белкам в особых участках плазмолеммы. называемых адгезионными соединениями, или фокальными контактами, которые связывают клетки друг с другом или клетки с компонентами межклеточного вещества
Актин - основной белок микрофиламентов Обычно молекула актина имеет вид двух спирально скрученных нитей (см. рис. 10-9 и 13-5). В микрофиламентах актин взаимодействует с рядом актин-связывающих белков (до нескольких десятков видов), выполняющих различные функции. Некоторые из них регулируют степень полимеризации актина, другие (например, филамин в кортикальной сети или фимбрин и виллин в микроворсинке) способствуют связыванию отдельных микрофиламентов в системы.Микрофиламенты более устойчивы к физическим и химическим воздействиям, чем микротрубочки.
Промежуточные филаменты - прочные и устойчивые в химическом отношении белковые нити толщиной около 10 нм (что является промежуточным значением между толщиной микротрубочек и микро-филаментов). Они встречаются в клетках разных тканей (см. ниже) и располагаются в виде трехмерных сетей в различных участках цитоплазмы, окружают ядро, входят в состав десмосом и полудесмосом эпителиальных клеток (в плазмолемме которых они закреплены посредством трансмембранных белков), лежат по всей длине отростков нейронов. Промежуточные филаменты образованы нитевидными белковыми молекулами, сплетенными друг с другом наподобие каната.
Функции промежуточных филаментов изучены недостаточно; установлено, однако, что они не влияют ни на движение, ни на деление клетки. К их основным функциям относятся:
(1) структурная - поддерживающая и опорная, обеспечение распределения органелл по определенным участкам цитоплазмы;
(2) обеспечение равномерного распределения сил деформации между клетками ткани, что препятствует повреждению отдельных клеток (благодаря связи промежуточных филаментов с трансмембранньгми белками десмосом и полудесмосом);
(3) участие в образовании рогового вещества в эпителии кожи; в эпителиальных клетках связываются с другими белками и образуют непроницаемые барьеры (роговые чешуйки), являются главным компонентом волос и ногтей;
(4) поддержание формы отростков нервных клеток и фиксация трансмембраниых белков (в частности, ионных каналов);
(5) удержание миофибрилл в мышечной ткани и прикрепление их к плазмолемме. что обеспечивает их сократительную функцию.
Очевидно, что функции, отмеченные цифрами (2)-(5), служат лишь частными проявлениями более общей структурной функции (1) в различных тканях.
Микротрубочки - наиболее крупные компоненты цитоскелета. Они представляют собой полые цилиндрические образования, имеющие форму трубочек, длиной до нескольких микрометров (в жгутиках более 50 им) диаметром около 24-25 нм, с толщиной стенки 5 нм и диаметром просвета 14-15 нм (рис. 3-14). 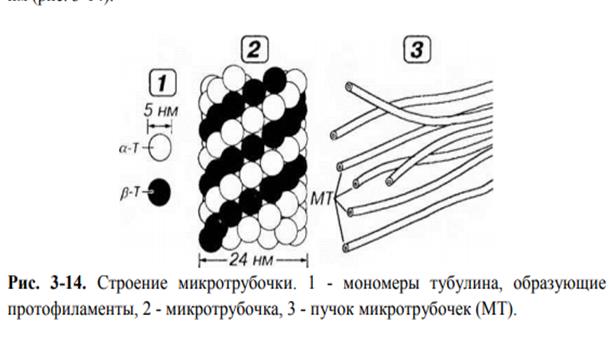
Стенка микротрубочки состоит из спиралевидно уложенных нитей - протофиламентов толщиной 5 нм (которым на поперечном разрезе соответствуют 13 субъединиц), образованных димерами из белковых молекул αи β-тубулина.
Функции микротрубочек: 1. поддержание формы и полярности клетки, распределения ее компонентов, 2. обеспечение внутриклеточного транспорта. 3. обеспечение движения ресничек, хромосом в митозе (формируют ахроматиновое веретено, необходимое для клеточного деления), 4. образование основы других органелл (центрнолей, ресничек).
Расположение микротрубочек. Микротрубочки располагаются в цитоплазме в составе нескольких систем: а) в виде отдельных элементов, разбросанных по всей цитоплазме и формирующих сети; б) в пучках, где они связаны тонкими поперечными мостиками (в отростках нейронов, в составе митотического веретена, манжетки сперматиды, периферического "кольца" тромбоцитов); в) частично сливаясь друг с другом с формированием пар, или дyблетов (в аксонеме ресничек и жгутиков), и триплетов (в базальном тельце и центриоли).
Угнетение самосборки микротрубочек посредством ряда веществ, являющихся ингибиторами митоза (колхицин, винбластин, винкристин), вызывает избирательную гибель быстроделящихся клеток. Поэтому некоторые из таких веществ успешно используются для химиотера- - 67 - паи опухолей. Блокаторы микротрубочек нарушают также транспортные процессы в цитоплазме, в частности, секрецию, аксонный транспорт в нейронах.
Разрушение микротрубочек приводит к изменениям формы клетки и дезорганизации ее структуры и распределения органелл
Дата добавления: 2021-03-18; просмотров: 273; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!