Роль молекул РНК в реализации наследственной информации
Лекция Экспрессия генов 31 октября 2017 г
Общая характеристика экспрессии генов.
Молекулярная организация генов про- и эукариот
Реализация генетической программы, заложенной в клетке, осуществляется частями в процессе транскрипции и трансляции информации, заключенной в соответствующих генах.
Ген представляет собой совокупность структурных и регуляторных полинуклеотидных последовательностей ДНК, определяющих возможность синтеза колинеарных молекул полипептида или РНК при данных условиях его экспрессии.
Ген состоит из двух оцДНК, одна из которых транскрибируется и называется кодогенной цепью (рис. 4.12).

Вторая цепь, комплементарная кодогенной цепи, получила название смысловой. Именно полинуклеотидная последовательность смысловой цепи используется для характеристики молекулярной организации гена и структуры генома.
Гены прокариот состоят из двух основных элементов: регуляторных и структурных последовательностей нуклеотидов (рис. 4.13).

Регуляторные последовательности контролируют инициацию, скорость, окончание транскрипции и другие функции. Регуляторные элементы располагаются как на 5΄-, так и на 3'-концах ДНК гена. К регуляторным элементам, локализованным на 5΄-конце гена, относятся промотор — последовательность нуклеотидов для связывания РНК-полимеразы, имеющаяся у всех генов, а также регуляторные последовательности, примыкающие к промотору, с которыми взаимодействуют регуляторные белки (репрессоры и активаторы), контролирующие скорость транскрипции. Указанные регуляторные последовательности имеются лишь у некоторых генов.
|
|
|
Промотор прокариот обычно содержит две канонические последовательности: область -35, или область узнавания, расположенную между нуклеотидами в положениях -30 и -35 от точки старта транскрипции, и область-10, или бокс Прибнова. Его последовательность локализуется между нуклеотидами -7 и -10. Знак «минус» указывает на то, что нуклеотиды находятся перед первым транскрибируемым нуклеотидом, которому соответствует положение +1. Обе эти последовательности специфически взаимодействуют с σ-фактором (сигма-фактором) РНК- полимеразы и играют большую роль в инициации транскрипции.
На З΄-конце гена локализуется регуляторная последовательность — терминатор, которая служит сигналом окончания транскрипции.
Структурные последовательности гена содержат информацию об аминокислотной последовательности поли- пептидной цепочки, или о первичной структуре тРНК, или рРНК и соответствуют транскриптону.
Около трети генов прокариот им оперонную организацию (рис. 4.14)
|
|
|
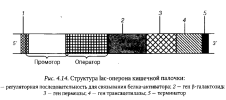
Опероном называют группу структурных генов, работа которых контролируется одним оператором — специфической полинуклеотидной последо- стельностью, частично входящей в состав промотора, с которой связываются регуляторные белки-репрессоры. Гены оперона имеют общий для них промотор.
Гены эукариот, так же как гены прокариот, состоят из двух основных элементов: регуляторных и структурных последовательностей нуклеотидов (рис. 4.15).

Отличительной чертой генов эукариот служит наличие у них дистальных регуляторных последовательностей, расположенных на расстоянии нескольких тысяч и даже сотен тысяч пар оснований от промотора, с которыми связываются регуляторные белки, получившие название специфических факторов транскрипции. Дистальные регуляторные последовательности для связывания транскрипционных факторов, активирующих и ускоряющих транскрипцию, называются энхансерами. Последовательности, с которыми связаются белки, блокирующие и тормозящие транскрипцию, получили название сайленсеров.
Структурные последовательности генов эукариот, в отличие от генов прокариот, состоят из информативных кодирующих (экзоны) и неинформативных некодирующих (интроны) участков ДНК.
|
|
|
Роль молекул РНК в реализации наследственной информации
Ген как участок молекулы ДНК, содержащий информацию о первичной структуре одной полипептидной цепи, непосредственного участия в синтезе белка не принимает. Роль посредника, функцией которого является перевод наследственной информации, сохраняемой в ДНК, в специфическую последовательность аминокислот в молекуле белка, играют рибонуклеиновые кислоты.
Рибонуклеиновые кислоты, в отличие от молекул ДНК, представлены одной полинуклеотидной цепью, которая построена из четырех видов нуклеотидов. Три из них, содержащие азотистые основания аденин, гуанин и цитозин, такие же, как в молекуле ДНК, и лишь один нуклеотид отличается: тимин в РНК замещен урацилом. Отдельные участки одноцепочечной РНК с комплементарными основаниями могут взаимодействовать друг с другом с образованием двухцепочечных фрагментов, в результате чего молекула РНК приобретает специфическую конформацию.
Основной функцией РНК является реализация программы синтеза белков, заложенной в ДНК клетки. Этот процесс состоит из нескольких этапов, каждый из которых осуществляется разными молекулами РНК. В соответствии с выполняемыми функциями различают несколько видов РНК.
|
|
|
Информационная РНК (иРНК), или матричная РНК (мРНК), — носитель генетической информации о первичной структуре белка. Содержится в ядре, цитоплазме, в митохондриях и пластидах. Ее второе название — матричная (мРНК) — объясняется тем, что в синтезе белка она играет роль, аналогичную типографской матрице. Молекулы мРНК содержат от 100 до нескольких тысяч нуклеотидов. На долю мРНК приходится до 5% общего содержания РНК в клетке. В связи с тем, что каждая отдельно взятая молекула мРНК содержит информацию о составе той или иной полипептидной цепи, количество разных мРНК в любой клетке очень велико. Все зрелые молекулы мРНК у эукариот имеют сходное строение и состоят из нескольких участков, выполняющих разные функции (рис. 4.16).
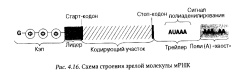
На 5'-конце зрелой молекулы мРНК находится «колпачок» (кэп) — участок, включающий один модифицированный нуклеотид. Этот участок всегда начинается 7-метилгуанозинтрифосфатом, который соединяется с соседним нуклеотидом пирофосфатной связью.
Колпачок защищает мРНК от разрушения их 5΄-экзонуклеазами. Непосредственно за колпачком располагается лидерный участок (5΄-нетранслируемый участок), длина которого обычно составляет от 40 до 80 нуклеотидов. В составе этого участка имеются нуклеотидные последовательности, комплементарные последовательностям рРНК малой субъединицы рибосомы. Указанные последовательности лидерного участка служат для связывания мРНК с рибосомой в процессе трансляции.
За лидерным участком следует инициирующий кодон АУГ, который кодирует у эукариот аминокислоту метионин (у прокариот — формилметионин). После завершения трансляции указанные аминокислоты обычно отщепляются от N-конца синтезированной полипептидной цепи. Инициирующий (АУГ) и терминирующий (УАА, УАГ и УГА) кодоны ограничивают кодирующий участок мРНК, в котором содержится информация о последовательности аминокислот в молекуле полипептида.
За кодирующим участком следует 3'-нетранслируемый участок, или трейлер, значительно превышающий по размеру лидерную область.
Непосредственно за трейлерным участком располагается поли(А)- фрагмент, или полиадениловый «хвост», состоящий из 150—200 адениловых нуклеотидов. Особенности строения трейлерного участка и поли(А)-фрагмента во многом определяют продолжительность функционирования зрелых молекул мРНК в клетках эукариот. Полиадениловый фрагмент по своей функции отчасти напоминает теломеры хромосомных ДНК: после завершения каждого раунда трансляции от полиаденилового «хвоста» молекулы мРНК отщепляется примерно 10—15 нуклеотидов. В гиалоплазме клеток молекулы мРНК обычно связаны с белками. Такие комплексы мРНК с белками называют информосомами.
Прокариотические мРНК имеют в основном такое же строение, как и мРНК эукариот. Отличия заключаются лишь в отсутствии у мРНК прокариот нетранслируемых участков, кэпа и полиаденилового «хвоста».
Кроме того, кодирующие участки прокариотических мРНК, траскриби- рующиеся на оперонах, обычно содержат несколько кодирующих участков — цистронов, которые отделены друг от друга неинформативными межцистрон- ными последовательностями.
Транспортная РНК (тРНК) имеет короткие цепи, состоящие из 70—100 нуклеотидов (рис. 4.17).

Она содержится в гилоплазме клетки, в митохондриях и пластидах. На 5΄-конце молекулы тРНК всегда находится гуанин, а на 3΄-конце — группировка из трех нуклеотидов ЦЦА. Все тРНК вследствие образования водородных связей между комплементарными основаниями разных участков цепи приобретают вторичную структуру, в двухмерном изображении напоминающую клеверный лист. В структуре тРНК выделяют акцепторный «стебель» и три петли — центральную (антикодоновую) и боковые. К 3΄-концу «стебля», заканчивающемуся последовательностью ЦЦА со свободной - ОН-группой, присоединяется транспортируемая аминокислота. На центральной петле тРНК располагается антикодон, состоящий из трех расположенных рядом нуклеотидов, которые комплементарны нуклеотидам соответствующего кодона мРНК.
При действии дополнительных водородных связей между нуклеотидами одноцепочечных петель образуется трехмерная третичная структура, похожая на перевернутую латинскую букву L.
В состав нуклеотидов тРНК входят метилированные основания. Они придают тРНК устойчивость к действию нуклеаз цитоплазмы и поддерживают третичную структуру молекулы.
В клетке имеется лишь 45 видов молекул тРНК, несущих разные антикодоны, а не 61, как можно было бы ожидать, исходя из количества кодонов мРНК, кодирующих аминокислоты. Это объясняется тем, что некоторые антикодоны тРНК могут связываться с несколькими кодонами мРНК. Функция тРНК состоит в переносе аминокислот к месту синтеза белка в рибосомах. На долю тРНК приходится примерно 15 % всей РНК, содержащейся в клетке.
Рибосомная РНК (рРНК) имеет самые крупные молекулы из всех видов РНК: в ее состав входит от 3 до 5 тыс. нуклеотидов. Рибосомная РНК синтезируется в ядрышке и участвует в образовании большой и малой субъединицы рибосом. В составе рибосом рРНК выполняет структурную и каталитическую функции. На долю рРНК приходится до 80 % всей РНК клетки.
В последние десятилетия открыты новые виды некодирующих молекул РНК, существенная особенность которых состоит в том, что большинство из них имеют небольшие размеры.
Малые ядерные РНК (мяРНК) — некодирующие молекулы РНК — встречаются только в ядрах эукариотических клеток. Они участвуют в процессах сплайсинга и поддержании целостности теломер.
МикроРНК, или миРНК, имеют размер 21-22 нуклеотида. У человека обнаружено более 700 разных видов миРНК. Молекулы миРНК играют большую роль в процессе трансфекции и регуляции экспрессии генов.
Маленькие интерферирующие РНК, или сиРНК, имеют длину 20-15 нуклеотидов. Выполняют сходные с миРНК функции и отличаются от них лишь своим биогенезом.
Транскрипция
Транскрипция - процесс перевода генетической информации в виде последовательности нуклеотидов в молекуле ДНК в последовательность нуклеотидов в молекуле мРНК. Иными словами, транскрипция — это процесс синтеза мРНК на матрице ДНК.
Транскрипция осуществляется с участием ряда ферментов, а также различных белковых факторов транскрипции (табл. 4.2; 4.3).
Таблица 4.2
Ферменты и белковые факторы,
Дата добавления: 2021-03-18; просмотров: 65; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!