Ядро. Деление ядра и клетки. Производные протопласта.
Лекция №1
Введение в ботанику. Цитология. Компоненты клетки. Цитоплазма.
Введение в ботанику.
Ботаника - наука о растениях. Она является разделом более общей науки – биологии, которая изучает все живые существа, населяющие Землю.
Ботанические знания зародились и быстро накапливались с практической деятельностью человека. Ботаника как наука, является древнейшей. Основоположниками ее были выдающиеся деятели древнего мира Аристотель (384 - 322 гг. до н. э.) и Феофраст (371 - 286 гг. до н. э.). Они обобщили накопленные сведения о разнообразии растений и их свойствах, приемах возделывания, размножения и использовании, географическом распространении.
В наши дни ботаника представляет собой большую многоотраслевую науку. Общая задача ее состоит в изучении отдельно взятых растений и их совокупностей - растительных сообществ. Структура и закономерности роста растений, их отношения с окружающей средой, закономерности распространения и распределения отдельных видов и всего растительного покрова на земном шаре; происхождение и эволюция царства растений, причины его разнообразия и классификация; запасы в природе хозяйственно ценных растений и пути их рационального использования, разработка научных основ введения в культуру (интродукции), лесоводственных, лекарственных, плодовых, овощных, технических и других растений - далеко не полный перечень вопросов, которые рассматриваются ботаническими науками.
|
|
|
Разделы ботаники.
Ботанику как часть более общей науки - биологии, в свою очередь, подразделяют на ряд частных наук, в задачи которых входит изучение тех или иных закономерностей строения и жизни растений или растительного покрова.
Анатомия – рассматривает внутреннее строение органов растений и слагающих их тканей.
Морфология – изучает форму растений, внешнее строение органов и их видоизменения.
Физиология растений – изучает процессы жизнедеятельности растений и закономерности превращения в них веществ.
Систематика - наука о разнообразии видов и причинах этого разнообразия. Задача систематики - приведение в легко обозримую научную систему всех наших знаний о видах. На основании целой серии методов систематик объединяет родственные виды в систематические группы более высокого ранга - роды, семейства и т. д.
Дендрология – рассматривает древесные растения, взаимоотношения их со средой.
География растений (фитогеография) - крупнейший раздел ботаники, основная задача которого состоит в изучении закономерностей распространения и распределения растений и их сообществ (ценозов) на суше и в воде.
|
|
|
Экология. Жизнь растений зависит от окружающей среды (климата, почвы и др.), но и растения, в свою очередь, влияют на создание этой среды - принимают участие в почвообразовательном процессе, изменяют климат. Задача экологии - изучение строения и жизни растений в связи с окружающей средой. Эта наука имеет первостепенное значение для практического земледелия.
Интродукция – изучает о переселении растений и введения их в культуру.
Микробиология - наука об особенностях жизненных процессов, происходящих в микроскопических организмах, преобладающую часть которых составляют бактерии и некоторые грибы. Успехи почвенной микробиологии широко используют в сельскохозяйственной практике.
Палеоботаника - наука об ископаемых растениях прошлых геологических периодов.
Другие разделы ботаники настолько обособились в связи с решением специальных задач и применяемыми методами работы, что давно уже составляют особые науки, например биофизика, биохимия, радиобиология, генетика и др.
Анатомия растений.
Анатомия (цитология растений) - наука о микроскопической и субмикроскопической структуре клетки и ее жизнедеятельности.
Клетка - это элементарная структурная и функциональная единица тела растений и животных, способная к самовоспроизведению. Первым увидел клетку английский естествоиспытатель Р. Гук при изучении покровной ткани бузины - пробки. Он усовершенствовал микроскоп, изобретенный Г. Галилеем в 1609 г., и использовал его для исследования тонких срезов органов растений. Свои наблюдения Р. Гук изложил в сочинении «Микрография», изданном в 1665 г., где он впервые применил термин «клетка». В 1833 г. английский ботаник Р. Броун обнаружил ядро, а в 1839 г. чешский физиолог Я. Пуркинье - цитоплазму. Многочисленные наблюдения клеточного строения растений и животных позволили немецким ученым - ботанику М. Шлейдену и зоологу Т. Шванну - в 1838 - 1839 гг. сформулировать клеточную теорию, суть которой заключается в том, что клетка - это основная элементарная структурная единица всех живых организмов. Клеточная теория доказывает единство происхождения, строения и эволюции растений и животных.
|
|
|
К концу XIX в. цитология окончательно сформировалась в самостоятельную науку. На базе светового микроскопа были изучены основные компоненты клетки, накапливались данные об их функциях.
Дальнейший прогресс цитологии связан с изобретением электронного микроскопа. В биологии его начали использовать в середине XX в.
|
|
|
Методы исследования клетки.
Применяемые для изучения клеток методы очень разнообразны. Основной из них - микроскопический. Большую роль продолжает играть световой микроскоп, современные модели которого дают увеличение до 2 тыс. раз. Но возможности светового микроскопа ограничены, частицы менее 0,2 мкм рассмотреть при помощи такого микроскопа невозможно. Электронный микроскоп дает увеличение в 200 - 300 тыс. раз и более. Здесь вместо пучка света используют поток электронов, движущихся с высокой скоростью.
Методом культуры тканей изучают структуру и жизнедеятельность живых клеток вне организма.
Цитохимический метод позволяет выявить наличие и определить количество различных веществ в клетке.
Разделить компоненты клетки с различной плотностью для изолированного изучения их можно с помощью метода центрифугирования. Извлечь из клетки отдельные компоненты (ядро, митохондрии и др.) позволяет метод микроскопической хирургии.
Разнообразие клеток.
Все разнообразие форм клеток можно свести к двум группам: паренхимные клетки - длина равна ширине или превышает ее не более чем в 2 - 3 раза; прозенхимные клетки - длина превышает ширину во много раз. Средняя длина клеток высших растений 10 - 100 мкм. Наиболее крупные паренхимные клетки достигают в длину несколько миллиметров и видны невооруженным глазом, например клетки плодов арбуза, лимона, клубней картофеля. Но особенно большую длину имеют прозенхимные клетки стеблей льна (40 мм), крапивы (80 мм), рами (200 мм).
Компоненты клетки.
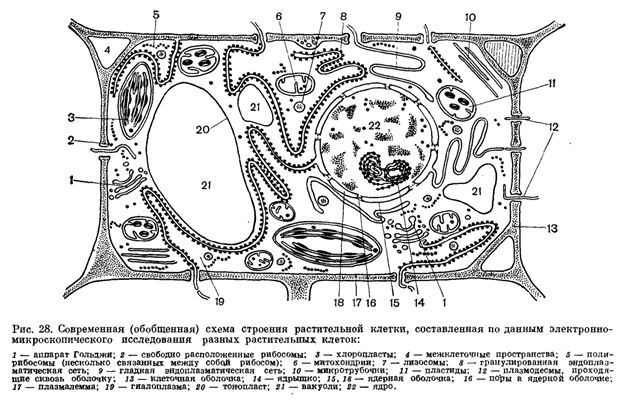
Рассматривая взрослую растительную клетку при помощи светового микроскопа, можно увидеть следующие компоненты: плотную стенку, одну большую или 2 - 3 небольшие вакуоли, занимающие центральную часть клетки; цитоплазму, расположенную между стенкой клетки и вакуолью; находящееся в цитоплазме ядро.
Ядро и цитоплазма - живые части клетки и в совокупности составляют протопласт. Стенка и вакуоли - неживые части клетки, производные протопласта, продукты его жизнедеятельности.
Цитоплазма - сложная структурная система, имеет мембранную организацию, состоит из гиалоплпзмы и органелл. Ее структуру образуют тонкие (4 - 10 нм), довольно плотные пленки - биологические мембраны. Основу их составляют липиды. Молекулы липидов расположены упорядоченно - перпендикулярно к поверхности, в два слоя, так, что их части, интенсивно взаимодействующие с водой (гидрофильные), направлены наружу, а части, инертные по отношению к воде (гидрофобные), - внутрь. Молекулы белка расположены несплошным слоем на поверхности липидного каркаса с обеих его сторон. Часть их погружена в липидный слой, а некоторые проходят через него насквозь, образуя участки, проницаемые для воды.
Мембраны образуют пограничный слой цитоплазмы, а также внешнюю границу ее органелл и участвуют в создании их внутренней структуры. Они делят цитоплазму на изолированные отсеки, в которых одновременно и независимо друг от друга могут протекать биохимические процессы часто в противоположном направлении (например, синтез и распад).
Одно из основных свойств биологических мембран - избирательная проницаемость (полупроницаемость): одни вещества проходят через них с трудом, другие легко и даже в сторону большей концентрации. Мембраны во многом определяют химический состав цитоплазмы и клеточного сока.
Плазмодесмы - тончайшие цитоплазматические нити, которые осуществляют связь между клетками. Клеточная пластинка, образующаяся при цитокинезе, пронизана трубочками эндоплазматического ретикулума, которые не разобщаются. На их основе и формируются плазмодесмы. Стенка плазмодесменного канала выстлана плазмалеммой, соединяющей плазмалеммы смежных клеток. В центре канала проходит трубка, сохраняющая непрерывность ретикулума обеих клеток. Между плазмалеммой и трубкой находится гиалоплазма, также непрерывная для обеих клеток. Плазмодесмы чаще всего бывают собраны в группы по нескольку десятков. Поодиночке они располагаются в стенках, не имеющих вторичных утолщений.
Плазмалемма и тонопласт. Плазмалемма - это мембрана, отграничивающая цитоплазму от стенки клетки и обычно плотно прилегающая к ней. Иногда плазмалемма бывает волнистой или образует глубокие складки. Она регулирует обмен веществ клетки с окружающей средой, а также участвует в синтезе веществ. Тонопласт отграничивает цитоплазму от вакуоли. Функция его та же, что и плазмалеммы.
Гиалоплазма. Это жидкая непрерывная среда, в которую погружены органеллы. Гиалоплазма содержит ферменты и нуклеиновые кислоты. Белки, входящие в состав гиалоплазмы, образуют сеть из тонких фибрилл (диаметром 2 - 3 нм) - трабекулярную систему, которая связывает между собой органеллы. Эта система очень динамична, она может распадаться при изменении внешних условий. Гиалоплазма способна к активному движению, которое может быть вращательным - вдоль стенки клетки, если в центре находится одна большая вакуоль, и струйчатым - по тяжам, пересекающим центральную вакуоль, в разных направлениях. Скорость движения зависит от температуры, интенсивности света, снабжения кислородом и других факторов. При движении гиалоплазма увлекает за собой органеллы. Гиалоплазма осуществляет взаимосвязь органелл, участвует в обмене, транспорте веществ, передаче раздражения и т. д.
Эндоплазматический ретикулум (эндоплазматическая сеть). Представляет собой отграниченную, мембранами систему взаимосвязанных субмикроскопических каналов и цистерн, пронизывающих гиалоплазму. Имеются две формы ретикулума: гранулярный (шероховатый) и агранулярный (гладкий). Гранулярный эндоплазматический ретикулум несет на поверхности мелкие гранулы - рибосомы. Агранулярный эндоплазматический ретикулум состоит из ветвящихся трубочек, отходящих от цистерн гранулярного ретикулума, не имеет рибосом. Обычно он развит слабее, чем гранулярный. Участвует в синтезе и транспорте эфирных масел, смол, каучука.
Рибосомы. Это гранулы диаметром около 20 нм, расположенные в гиалоплазме или прикрепленные к поверхности мембран эндоплазматического ретикулума. Они обнаружены также в митохондриях и пластидах. Рибосомы состоят из белка и рибонуклеиновой кислоты (РНК) и не имеют мембранной структуры. Функция рибосом - синтез белка, самовоспроизводство живой материи. Этот процесс происходит в рибосомах, расположенных группой и связанных между собой нитевидной молекулой и-РНК. Такие группы называют полисомами. Считают, что рибосомы формируются в ядре.
Аппарат Гольджи. Состоит из диктиосомы и пузырьков Гольджи. Диктиосома представляет собой стопку из 5 - 7 плоских цистерн, ограниченных агранулярной мембраной. Диаметр цистерн около 1 мкм, толщина 20 - 40 нм. Цистерны не соприкасаются друг с другом. Пузырьки Гольджи отчленяются от краев цистерн и распространяются по всей гиалоплазме. В диктиосоме происходят синтез, накопление и выделение полисахаридов. Пузырьки Гольджи транспортируют их, в том числе и к плазмалемме. Мембрана пузырьков встраивается в плазмалемму, а содержимое оказывается снаружи от плазмалеммы и может включаться в стенку. Пузырьки Гольджи могут включаться и в тонопласт.
Происхождение диктиосом еще точно не установлено. Считают, что в их образовании принимает участие эндоплазматический ретикулум. В некоторых клетках аппарат Гольджи отсутствует.
Сферосомы. Округлые блестящие тельца диаметром 0,5 - 1 мкм. Это центры синтеза и накопления растительных масел. Они отшнуровываются от концов тяжей эндоплазматического ретикулума. Мембрана, расположенная на поверхности сферосомы, по мере накопления масла редуцируется, и от нее остается только наружный слой.
Лизосомы. Пузырьки размером 0,5-2 мкм, имеющие на поверхности мембрану. Содержат ферменты, которые могут расщеплять белки, липиды, полисахариды и другие органические соединения. Образуются так же, как и сферосомы, из тяжей эндоплазматического ретикулума. Их функция - разрушение отдельных органелл или участков цитоплазмы (локальный автолиз), необходимое для обновления клетки.
Митохондрии. Форма митохондрий чрезвычайно разнообразна - овальная, округлая, цилиндрическая, гантелевидная, ветвистая и т.д. Длина их равна 2 - 5 мкм, диаметр - 0,3 - 1 мкм. На поверхности митохондрии находятся две мембраны. Внутренняя мембрана образует выросты в полость митохондрии в виде гребней или трубочек, называемых кристами. Кристы значительно увеличивают мембранную поверхность митохондрии. Пространство между кристами заполнено жидким веществом - матриксом, в котором находятся рибосомы и содержится дезоксирибонуклеиновая кислота (ДНК). Поверхность внутренней мембраны покрыта мельчайшими тельцами, имеющими шаровидную головку и ножку (АТФ-сомы).
Митохондрии - это энергетические лаборатории клетки. Здесь происходят расщепление углеводов, жиров и других органических веществ при участии кислорода (дыхание) и синтез АТФ. Выделяемая при дыхании энергия преобразуется в энергию макроэргических связей молекулы АТФ, которая затем используется для осуществления процессов жизнедеятельности клетки - деления, поглощения и выделения веществ, синтеза и т.д.
Пластиды. Бывают только у растений. Эти органеллы имеют на поверхности две мембраны. В зависимости от окраски различают три типа пластид: хлоропласт - зеленого цвета; хромопласты - желтого, оранжевого, красного цветов; лейкопласты - бесцветные.
Хлоропласты содержат зеленый пигмент хлорофилл, а также пигменты из группы каротиноидов - каротин (оранжевый) и ксантофилл (желтый). Именно с хлорофиллом связана основная функция хлоропластов - синтез органических веществ из неорганических при участии энергии света (фотосинтез). Поэтому хлоропласты присутствуют только в клетках наземных органов, на которые падает солнечный свет. Хлоропластам растения обязаны зеленой окраской.
У высших растений хлоропласты в большинстве случаев имеют линзовидную форму. Диаметр их 4 - 6 мкм, толщина 1 - 3 мкм. У водорослей хлоропласты, называемые также хроматофорами, очень разнообразны по форме и величине. Они могут иметь звездчатую форму, лентовидную, сетчатую и др. Обычно в клетке имеются от 1 до 50 хлоропластов. Располагаются они в постенном слое цитоплазмы.
Внутри хлоропластов находится однородное вещество - строма, пронизанная системой параллельно расположенных мембран. Мембраны имеют вид плоских мешков, их называют тилакоидами, или ламеллами. У большинства высших растений часть тилакоидов имеет дисковидную форму. Эти тилакоиды собраны в стопки, называемые гранами. Хлорофилл и каротиноиды находятся в каждой из двух мембран тилакоида граны. Граны связаны между собой тилакоидами стромы. Внутренняя мембрана оболочки хлоропласта иногда образует складки и переходит в тилакоиды стромы. В строме находятся молекулы ДНК, рибосомы, капли липидов, называемые пластоглобулами, крахмальные зерна и другие включения.
Крахмал образуется в хлоропластах из продуктов фотосинтеза, его называют фотосинтетическим, или первичным. При помощи ферментов фотосинтетический крахмал осахаривается и в виде глюкозы транспортируется из листа на построение органов или в запас.
Лейкопласты не содержат пигментов. По размеру они значительно меньше хлоропластов и не имеют строго определенной формы. Лейкопласты присутствуют чаще всего в клетках тканей и органов, на которые не падает солнечный свет: в корнях, клубнях, семенах и др. Внутренняя мембранная система у лейкопластов развита значительно слабее, чем у хлоропластов. В строме имеются молекулы ДНК, рибосомы, пластоглобулы. Основная функция лейкопластов - синтез и накопление запасных питательных продуктов, в первую очередь крахмала, иногда белков, редко масла. Лейкопласты, накапливающие крахмал, называют амилопластами. В них из сахаров, поступающих из фотосинтезирующих органов, образуются крахмальные зерна различного размера и формы - вторичный крахмал. Запасной белок может откладываться в виде кристаллов или аморфных гранул, масло - в виде пластоглобул.
Хромопласты содержат красные, оранжевые, желтые пигменты из группы каротиноидов. По размеру они меньше хлоропластов, форма их очень разнообразна. Внутренняя мембранная система у них чаще всего отсутствует.
В зависимости от формы накопления каротиноидов различают хромопласты глобулярного, фибриллярного (трубчатого) и кристаллического типов. У хромопластов наиболее распространенного глобулярного типа пигменты растворены в пластоглобулах. У хромопластов фибриллярного типа каротиноиды не только присутствуют в пластоглобулах, но и образуют группы параллельных нитей или трубок, расположенных в строме. У хромопластов кристаллического типа каротиноиды присутствуют главным образом в виде кристаллов различной формы, определяющих форму самой пластиды (серповидную, ромбовидную, игловидную и т. д.).
Хромопласты встречают в клетках лепестков некоторых растений, зрелых плодов, осенних листьев. Их функция в процессе обмена веществ не выяснена. Косвенное биологическое значение хромопластов состоит в привлечении насекомых для перекрестного опыления и животных для распространения семян.
В процессе эволюции первыми из пластид появились хлоропласты, из которых при расчленении тела растения на органы образовались лейкопласты и хромопласты. В онтогенезе почти все виды пластид могут переходить друг в друга. Наиболее часто происходит превращение лейкопластов в хлоропласты (например, при образовании зародыша из оплодотворенной яйцеклетки) и хлоропластов в хромопласты (например, при осеннем пожелтении листьев). Лишь хромопласты в природных условиях, как правило, не превращаются в другие виды пластид. Количество пластид в клетке увеличивается за счет деления их путем перетяжки.
Ядро. Деление ядра и клетки. Производные протопласта.
Ядро.
Ядро может функционировать только в цитоплазматической среде. Это - место хранения и воспроизводства наследственной информации, определяющей признаки данной клетки и всего организма в целом, а также центр управления синтезом белка. Если из клетки удалить ядро, то она вскоре погибнет. Обычно в клетке имеется одно ядро, но у некоторых видов водорослей и у грибов многоядерные клетки. Бактерии и сине-зеленые водоросли не имеют оформленного ядра.
Форма ядра разнообразна, но обычно соответствует форме клетки: в паренхимных клетках чаще всего шаровидная, в прозенхимных - линзовидная или веретеновидная. Диаметр ядра клеток вегетативных органов покрытосеменных растений 10 - 25 мкм. У плесневых грибов диаметр ядра всего 1 - 2 мкм, а у харовых водорослей может достигать 2,5 мм. В процессе онтогенеза форма, размер и местоположение ядра в клетке могут изменяться.
Под световым микроскопом ядро имеет вид пузырька с 1 -3 темными пятнышками - ядрышками. Оно состоит из ядерной оболочки, нуклеоплазмы, хромосом, ядрышек.
Ядерная оболочка отграничивает содержимое ядра от цитоплазмы. Состоит из двух мембран с промежутком между ними, называемым перинуклеарным пространством. Толщина мембран 10 нм, а толщина перинуклеарного пространства варьирует. Общая толщина оболочки 40 - 80 нм. Внутренняя мембрана оболочки агранулярная, к наружной мембране прикреплены рибосомы. По структуре и химическому составу ядерная оболочка близка к эндоплазматическому ретикулуму, тем более что ее наружная мембрана образует выросты, переходящие в ретикулум цитоплазмы. Ядерная оболочка имеет особые образования - ядерные поры. Это сложные структуры. По границе поры, образованной в результате слияния двух мембран, расположены гранулы, от которых отходят фибриллы. Часть фибрилл сходится в центре, формируя диафрагму. Диаметр поры 80-90 нм. Через поры макромолекулы проходят из нуклеоплазмы в гиалонлазму и в обратном направлении. Ядерная оболочка контролирует обмен веществ между ядром и цитоплазмой, способна к синтезу белков и липидов.
Нуклеоплазма представляет собой коллоидный раствор, в котором размещены хромосомы и ядрышки. В состав нуклеоплазмы входят различные ферменты, нуклеиновые кислоты. Она не только осуществляет связь между органеллами ядра, но и трансформирует вещества, проходящие через нее.
Хромосомы могут находиться в двух состояниях, В рабочем состоянии это деконденсированные в различной степени, тонкие (10 нм) нитчатые структуры, активно участвующие в процессе обмена веществ. Они видны только под электронным микроскопом. Во время деления ядра хромосомы максимально конденсируются, становятся короткими и толстыми (видны под световым микроскопом). Выполняют функцию распределения и переноса генетической информации, в процессе обмена веществ не участвуют, поглощают многие красители и интенсивно окрашиваются.
По химической природе хромосома представляет собой нуклеопротеид, состоящий из ДНК и белка. Составные части (мономеры) молекулы ДНК - нуклеотиды. Нуклеотид имеет три компонента - остаток фосфорной кислоты, сахар дезоксирибозу и одно из четырех азотистых оснований: аденин, гуанин, тимин, цитозин. Нуклеотиды соединяются в длинную цепь. Молекула ДНК состоит из двух таких чрезвычайно длинных цепей, которые связаны между собой азотистыми основаниями, причем аденин всегда соединяется с тимином, а гуанин - с цитозином. Эта двойная цепь закручена вокруг оси. Одно из важнейших свойств молекулы ДНК - репликация (самоудвоение), при которой цепочки нуклеотидов расходятся и каждая из них достраивает утраченную. Участок молекулы ДНК, определяющий способность к синтезу одной полипептидной цепи, называют геном. Белок в хромосоме располагается на поверхности молекулы ДНК в виде футляра.
Хромосома имеет первичную перетяжку (неконденсированный участок), где находится центромера (пластинчатая структура дисковидной формы), а иногда и вторичную, которая отделяет от нее небольшой фрагмент - спутник.
Каждый вид растений содержит в клетке строго определенное число хромосом. В соматических клетках это число обычно диплоидное (2n). Оно образуется в результате слияния двух половых клеток с гаплоидным (моноплоидным) числом хромосом (n).
Ядрышко. Обычно это сферическое тельце диаметром 1 - 3 мкм, состоящее в основном из белка и РНК. Молекула РНК, как и молекула ДНК, представляет цепочку нуклеотидов, но нуклеотид РНК содержит вместо дезоксирибозы рибозу, а вместо тимина урацил. В отличие от молекулы ДНК, молекула РНК имеет лишь одну такую цепочку.
Ядрышко обычно контактирует со вторичной перетяжкой хромосомы, называемой организатором ядрышка, на которой происходит матричный синтез р-РНК. Затем р-РНК объединяется с белком, в результате образуются гранулы рибонуклеопротеидов - предшественников рибосом, которые попадают в нуклеоплазму и через поры ядерной оболочки проникают в цитоплазму, где заканчивается их оформление.
В ядре может быть одно или несколько ядрышек.
Деление ядра и клетки.
В процессе онтогенеза ядро возникает только от ядра. Деление ядра всегда предшествует делению клетки.
Существует три способа деления клетки: митоз, амитоз, мейоз.
Митоз - наиболее универсальный способ деления соматических клеток. Он свойствен организмам всех уровней эволюционного развития, имеющим оформленное ядро. Совокупность саморегулирующихся процессов, происходящих в клетке от одного деления до другого, называют митотическим циклом. Митотический цикл состоит из интерфазы и митоза, тесно связанных между собой; продолжительность его примерно 15 - 30 ч.
Интерфаза - наиболее продолжительная часть митотического цикла. Ядро относительно крупное, с хорошо заметными 1 - 2 ядрышками и слабозернистой структурой. Хромосомы деконденсированы, и окраска не выявляет их. В этой фазе происходят важные биохимические процессы, подготавливающие клетку к делению. В интерфазе различают три периода: пресинтетический (G1) - происходят рост вновь образовавшейся клетки, воссоздание цитоплазматических структур, синтез РНК и белка, накопление энергии; синтетический (S) - репликация молекул ДНК, образование двух хроматид; постсинтетический (G2) - синтез белка, накопление энергии.
Митоз делят на четыре фазы: профазу, метафазу, анафазу и телофазу. Продолжительность митоза 1 - 3 ч.
В начале профазы ядро увеличивается, и в нем отчетливо видны спутанные в клубок хромосомы, начавшие конденсироваться. К концу профазы хромосомы укорачиваются. Иногда заметно, что они состоят из двух хроматид. Ядрышко к этому моменту обычно дезинтегрируется. Ядерная оболочка распадается на небольшие фрагменты, неотличимые от элементов эндоплазматического ретикулума. Нуклеоплазма смешивается с гиалоплазмой. На полюсах клетки появляются белковые нити, растущие к центру. Профаза - самая продолжительная фаза митоза.
В начале метафазы хромосомы достигают максимальной конденсации и передвигаются к экваториальной пластинке клетки. Они хорошо видны в оптический микроскоп. Число, форма, размер и расположение митотических хромосом характерны и постоянны для каждого вида растения (кариотип). Их графическое изображение составляет идиограмму вида. В диплоидном наборе имеются пары одинаковых по структуре хромосом, называемых гомологичными. Соответственно гаплоидный набор содержит по одной из гомологичных хромосом.
В метафазе хроматиды отделяются друг от друга, связь между ними сохраняется только в центромере. Из нитей формируется веретено деления. Оно состоит из опорных и тянущих нитей. Опорные нити идут от одного полюса клетки к другому через экваториальную пластинку, а тянущие связывают центромеры хромосом с полюсами. Нити веретена деления не всегда видны, так как ядерный краситель не окрашивает их. Наиболее характерно для метафазы то, что центромеры хромосом, прикрепленные к нитям веретена, располагаются в плоскости экваториальной пластинки клетки.
В анафазе центромера разделяется и хроматиды расходятся к полюсам вследствие сокращения тянущих нитей. Каждая хроматида становится хромосомой. Следовательно, на каждом полюсе оказывается столько хромосом, сколько их было у исходной клетки. Анафаза - самая короткая фаза.
В телофазе происходит процесс, противоположный происходящему в профазе: хромосомы деконденсируются, веретено деления разрушается, образуются ядерная оболочка и ядрышки. В начале телофазы хромосомы - в виде двух темных сгустков на полюсах клетки, к концу их контур исчезает. В это же время в экваториальной пластинке клетки появляются волокна, располагающиеся перпендикулярно к ней, - фрагмопласт. В центре фрагмопласта накапливаются пузырьки Гольджи, содержащие пектиновые вещества. Они дают начало клеточной пластинке, которая, разрастаясь центробежно и раздвигая фрагмопласт, достигает стенок материнской клетки. Происходит цитокинез, завершающий митоз. Позднее на клеточной пластинке с обеих сторон формируются первичные стенки. В результате митотического цикла образуются две клетки, имеющие хромосомы, идентичные хромосомам материнской клетки по структуре ДНК, форме, размеру и числу, то есть обеспечивается наследственное сходство дочерних клеток с исходной материнской.
Амитоз - это другой способ деления соматических клеток. Сущность его состоит в том, что ядро делится на две или более частей без каких-либо предшествующих изменений структуры. Вслед за перешнуровкой ядра следует деление цитоплазмы. При амитозе хромосомы распределяются между дочерними клетками неравномерно, поэтому не обеспечивается их биологическая равноценность. Но образовавшиеся клетки не теряют своей структурной организации. Амитоз наблюдают в клетках высокодифференцированных и старых тканей.
Мейоз столь же универсален, как и митоз, но он свойствен небольшой группе клеток, возникающих в связи с размножением. Сущность мейоза состоит в сокращении числа хромосом в два раза. Биологический смысл этого процесса раскрыт уже давно. Дело в том, что гаплоидные половые клетки - гаметы (n) при половом процессе сливаются и образуют зиготу. При этом число хромосом увеличивается в два раза (2n), а зигота получает наследственную информацию от обеих гамет. Следовательно, благодаря мейозу сохраняется постоянство числа хромосом. Мейоз состоит из двух делений, неизменно следующих одно за другим. В каждом из них выделяют те же четыре фазы, что и в митозе, но они имеют принципиальные различия. Первое деление сложное и специфическое, сопровождается сокращением числа хромосом. В профазе этого деления между гомологичными хромосомами происходит обмен участками (кроссинговер). В анафазе к полюсам клетки расходятся не хроматиды, как при митозе, а гомологичные хромосомы. Второе деление осуществляется по типу митоза. В результате из одной диплоидной клетки получаются четыре (тетрада) дочерних клетки с гаплоидным числом хромосом. Разнообразие их хромосомных наборов обусловливает разнообразие признаков у последующих поколений. Это - основа для эволюции вида
В некоторых случаях образованию половых клеток не предшествует сокращение числа их хромосом, и они остаются диплоидными. В результате образующееся из зиготы растение будет содержать 3 - 4 набора хромосом, а иногда и более. Такие клетки и состоящие из них растения называют полиплоидными. Полиплоидные растения обычно имеют более крупные размеры. Многие высокопродуктивные сорта томата, кукурузы, пшеницы и других возделываемых растений - полиплоиды. Полиплоидию можно вызвать искусственно.
Производные протопласта.
Большинство продуктов жизнедеятельности протопласта включено в вакуоли и цитоплазму. Некоторые, например клеточная стенка, откладываются вне протопласта, образуя скелетную основу клетки. Эргастические вещества подразделяют на физиологически активные, запасные, экскреторные, входящие в состав стенки клетки.
Вакуоль .
Это полость, заполненная жидким содержимым - клеточным соком, ограниченная тонопластом. Образуются вакуоли из локальных расширений эндоплазматического ретикулума, в которых накапливается клеточный сок. Эти расширения отчленяются от ретикулума и округляются, а мембрана ретикулума становится тонопластом. В образовании вакуолей могут, по-видимому, участвовать и элементы аппарата Гольджи. Небольшие вакуоли имеются даже в очень молодых клетках. По мере роста последних объем вакуолей увеличивается. У большинства зрелых клеток имеется одна большая вакуоль, занимающая центральную часть клетки, и много мелких - рассеянных в постенном слое цитоплазмы. Если же ядро располагается в центре клетки, то цитоплазма, окружающая его, связана с постенным слоем тяжами, разделяющими центральную вакуоль на несколько более мелких.
Клеточный сок представляет собой водный раствор разнообразных органических и неорганических соединений, выделяемых протопластом. У разных видов растений и даже в разных органах одного растения химический состав клеточного сока неодинаков. Реакция клеточного сока обычно слабокислая или нейтральная.
Химический состав клеточного сока. Органические вещества: азотистые: белки (протеины, протеиды), аминокислоты (аспарагин, тирозин, лейцин и др.), алкалоиды (хинин, морфин, никотин, колхицин, кофеин и др.); безазотистые: углеводы (моносахариды - глюкоза, фруктоза; дисахарпды - сахароза, мальтоза; полисахариды - инулин), гликозиды (амигдалин, сапонин, соланин, пигменты - антоциан, антохлор и др.), дубильные вещества (танниды), органические кислоты (щавелевая, яблочная, винная, лимонная и др.), кристаллы (соли щавелевой и других кислот), эфирное масло и др. Неорганические вещества: нитраты, фосфаты, хлориды. Часть этих веществ, например углеводы, - запасные, другие - экскреторные, конечные продукты обмена веществ.
Оформленные компоненты клеточного сока представлены кристаллами оксалата кальция (CaC2О4). Щавелевая кислота - один из вредных продуктов обмена веществ в клетках. Растение освобождается от нее при помощи ионов кальция. Оксалат кальция откладывается в растениях главным образом в отмирающих клетках в виде одиночных кристаллов разнообразной формы: сросшихся - друз, собранных в пачку - рафид и др.. Особенно много кристаллов оксалата кальция образуется в органах, которые время от времени сбрасываются: в коре деревьев, листьях, сухих чешуях луковиц и др. Как правило, друзы бывают у двудольных растений, а рафиды - у однодольных.
Запасные продукты.
Запасные продукты - это вещества, временно выключенные из обмена веществ. Они расходуются на построение тела растения или на различные жизненные процессы как энергетический материал. Местом отложения их служат вакуоли и цитоплазма. В вакуолях запасные продукты накапливаются в виде растворов, в цитоплазме - в виде оформленных включений: алейроновых, крахмальных зерен, капель жирного масла и др.
Алейроновые зерна - это гранулы запасного белка. Они обычно образуются в клетках запасающей ткани зрелых семян. При формировании семян в мелких вакуолях накапливается белок. В созревающих семенах вакуоли теряют воду и превращаются в алейроновые зерна. При прорастании семян, когда они обогащаются водой, алейроновые зерна вновь преобразуются в вакуоли.
Алейроновые зерна имеют округлую форму, диаметр их колеблется от 0,2 до 20 мкм. Снаружи они покрыты мембраной. У простых алейроновых зерен белок находится в виде аморфной массы (бобовые, кукуруза, рис), у сложных - в аморфную массу включен один, реже 2 - 3 белковых кристалла и небольшое округлое тельце - глобоид, содержащее запасной фосфор
Белковые тела могут образовываться и в других частях клетки - в ядре, пластидах, митохондриях, эндоплазматическом ретикулуме.
Крахмальные зерна. Наиболее распространенный и важный вид запасных продуктов растений. Встречаются в клетках всех органов, но особенно богаты ими семена и подземные видоизмененные побеги (клубни, луковицы, корневища). Крахмальные зерна образуются только в пластидах. В хлоропластах откладываются зерна первичного крахмала. Но здесь он не накапливается. При помощи ферментов первичный крахмал осахаривается и в виде глюкозы транспортируется из листа в другие органы. Вторичное превращение сахара в крахмал происходит в лейкопластах (амилопластах). Образование зерен вторичного крахмала начинается в определенных точках стромы амилопласта, называемых образовательными центрами. Рост зерен идет путем наложения слоев. Смежные слои могут иметь различные показатели преломления и поэтому видны под микроскопом. Слоистость бывает концентрической и эксцентрической. С ростом крахмального зерна объем стромы амилопласта уменьшается, и в определенный момент слой ее становится настолько тонким, что неразличим под световым микроскопом. Надо помнить, что двухмембранная оболочка и тонкий слой стромы всегда присутствуют на поверхности зерна вторичного крахмала.
Если в амилопласте имеется один образовательный центр, то формируется простое зерно, если два и более - сложное зерно, состоящее как бы из нескольких простых. Полусложное зерно образуется в том случае, если крахмал сначала откладывается вокруг нескольких образовательных центров, а затем после соприкосновения простых зерен вокруг них возникают общие слои. Размер крахмальных зерен колеблется в больших пределах. Так, у картофеля диаметр их достигает 100 мкм, у пшеницы и ржи бывают мелкие зерна диаметром 2 - 9 мкм и крупные диаметром 30 - 45 мкм, у кукурузы - диаметром 5 - 30 мкм.
Форма, размер, структура крахмальных зерен специфичны для каждого вида растения, а иногда и для сорта. Это обстоятельство используют при анализе состава муки.
Капли жирного масла имеют сферическую форму, накапливаются в гиалоплазме. Число и размер их в клетках колеблются. Капли жирного масла содержатся в клетках всех органов, но наиболее богаты ими семена и плоды.
Стенка клетки.
Клетки растений, в отличие от клеток животных, имеют хорошо развитую, обычно твердую стенку. Совокупность клеточных стенок создает скелет растения, обладающий механической прочностью. Стенка играет важную роль в поглощении и передвижении веществ. Она часто сохраняется дольше, чем протопласт, поэтому и мертвые клетки продолжают выполнять разнообразные функции.
Первичная стенка образуется при делении клетки. К концу телофазы в экваториальной плоскости клетки возникает клеточная пластинка, впоследствии превращающаяся в срединную пластинку. Она состоит в основном из пектиновых веществ. На клеточную пластинку протопласт каждой из дочерних клеток откладывает первичную стенку. Толщина ее равна 0,1 - 0,5 мкм. В состав первичной стенки входят в основном пектиновые вещества, гемицеллюлозы и целлюлоза (всего 10-12 %), а также много воды.
Построение стенки и ее рост связаны с деятельностью аппарата Гольджи и плазмалеммы. Рост первичной стенки происходит путем внедрения молекул целлюлозы, синтезируемых на наружной поверхности плазмалеммы, и аморфных пектиновых веществ, доставляемых пузырьками Гольджи. При этом поверхность первичной стенки увеличивается, а толщина остается постоянной. Увеличение поверхности первичной стенки продолжается до достижения клеткой постоянного размера.
Вторичная стенка образуется путем наложения изнутри на первичную стенку новых слоев. При этом происходит рост стенки в толщину, а объем полости клетки уменьшается. Толщина вторичной стенки зависит от специализации клетки и колеблется от 1 до 10 мкм. Наиболее мощная она у клеток, выполняющих механическую функцию. У клеток, по которым передвигается вода, вторичная стенка откладывается в виде колец, спирали или имеет неравномерную толщину.
Вторичная стенка в основном состоит из целлюлозы (до 90%) и гемицеллюлоз. Пектиновых веществ и воды в ней значительно меньше, чем в первичной стенке.
Параллельное расположение молекул целлюлозы создает тонкую упорядоченность внутренней структуры стенки. Несколько десятков нитевидных молекул объединяются в мицеллы, из которых формируются микрофибриллы - основные структурные единицы стенки. Диаметр их равен 10 - 30 нм, а длина достигает нескольких микрометров. Микрофибриллы могут объединяться в фибриллы - волокна или пластинки толщиной 0,4 - 0,5 мкм, видимые в световой микроскоп. В первичных стенках паренхимных клеток микрофибриллы расположены беспорядочно. В первичных стенках прозенхимных клеток и во вторичных стенках микрофибриллы располагаются или параллельно продольной оси клетки (волокнистая текстура), или перпендикулярно к ней (кольчатая текстура), или под некоторым углом (спиральная текстура). В процессе роста стенки ориентация микрофибрилл может меняться, что обусловливает слоистость стенки. Между микрофибриллами находится жидкий матрикс, состоящий из воды, пектиновых веществ и гемицеллюлоз.
Стенки, состоящие из целлюлозы, эластичны и очень прочны. Однако в процессе жизнедеятельности в связи со специфической функцией химический состав стенки клетки может меняться, что вызывает изменение ее физических свойств. Часто в матриксе вторичной и первичной стенок и в срединной пластинке накапливается лигнин. Это ведет к потере эластичности, повышению твердости, снижению проницаемости. Такие стенки называют одревесневшими. Они выполняют скелетную (механическую) функцию. Стенки некоторых клеток содержат липиды - воск, кутин, суберин. Эти вещества не смешиваются с матриксом стенки, а образуют самостоятельные слои. Обычно кутин и суберин откладываются вместе с воском в виде чередующихся параллельных слоев: кутин - на наружной поверхности клеточных стенок, граничащих с внешней средой (кутикула), суберин - на внутренней поверхности клеточных стенок, граничащих с плазмалеммой. Воск может и один образовывать слои на наружной поверхности стенок. Липиды уменьшают испарение с поверхности клеток. Суберин непроницаем для воды и газов, поэтому после его отложения протопласт отмирает. Стенки, содержащие суберин, называют опробковевшими. В матриксе клетки могут накапливаться минеральные вещества в виде кристаллов. Чаще всего это кремнезем или соли кальция. Они придают стенке твердость и хрупкость. Растения, имеющие клетки с такими стенками, плохо поедаются животными.
Срединная пластинка соединяет клетки между собой. Разрушение срединной пластинки и разъединение клеток называют мацерацией. Она происходит в естественных условиях при перезревании плодов, в черешках листьев перед их опадением и т.д. При частичной мацерации, когда срединная пластинка разрушается только по углам клеток, происходит образование межклетников.
Поры - это места, где не образуется вторичная стенка. Они имеют вид каналов, идущих от полости клетки до первичной стенки. По форме канала различают поры простые и окаймленные. У простых пор канал на всем протяжении имеет примерно одинаковый диаметр. С поверхности они имеют вид кружков. У окаймленных пор канал по направлению к первичной стенке расширяется. Поэтому с поверхности они выглядят, как две концентрические окружности. Обычно в двух смежных клетках поры образуются друг против друга и имеют вид канала, разделенного тонкой перегородкой из срединной пластинки и двух первичных стенок, называемой замыкающей пленкой. У окаймленных пор замыкающая пленка в центральной части имеет утолщение (торус). В живых клетках замыкающие пленки пронизаны многочисленными плазмодесмами. Поры, как и плазмодесмы, облегчают транспорт веществ между клетками.
В стенках клеток, специализирующихся на передвижении веществ, под действием ферментов образуются крупные отверстия, называемые перфорациями.
Дата добавления: 2020-12-22; просмотров: 89; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!