Дыхание в условиях пониженного и повышенного атмосферного давления
Физиология дыхания
Главным условием существования животных является наличие в окружающей среде кислорода, необходимого для окислительного процесса, благодаря которому клетки получают энергию, используемую для основных проявлений жизни — ассимиляции и диссимиляции. В результате жизнедеятельности организма образуются различные продукты метаболизма, важнейшим из которых является диоксид углерода (СO2). При этом нормальная жизнедеятельность клетки возможна только при условии удаления этих продуктов из организма. Обмен кислорода и диоксида углерода между окружающей средой и тканями — одно из главных условий жизни организма.
Дыхание — совокупность процессов, в результате которых происходит поступление кислорода в организм и выделение из него углекислого газа.
Функциональная система дыхания — совокупность структур, которые обеспечивают необходимый объем легочной вентиляции (внешнее звено саморегуляции), поддерживая оптимальный для метаболизма уровень pO2, pCO2 и рН крови и тканей (внутреннее звено саморегуляции).
Нормальное функционирование организма животных возможно только при условии пополнения энергией, которая непрерывно расходуется. Организм получает энергию за счет окисления органических веществ — белков, жиров, углеводов. При этом высвобождается скрытая химическая энергия, которая служит источником жизнедеятельности, развития и роста организма. Таким образом, значение дыхания состоит в поддержании в организме оптимального уровня окислительно-восстановительных процессов.
|
|
|
Этапы дыхания
Комплекс последовательных физиологических и физико-химических процессов, обеспечивающих дыхание, подразделяют на пять этапов.
1-й этап — внешнее дыхание, или вентиляция легких — процессы, обеспечивающие ритмическое поступление определенных объемов атмосферного воздуха в легкие (вдох) и удаление его из легких в атмосферу (выдох).
2-й этап — диффузия газов в легких (газообмен в легких) — процессы, обеспечивающие переход кислорода из альвеолярного воздуха в кровь и углекислого газа в обратном направлении.
3-й этап — транспорт газов кровью — процессы, обеспечивающие растворение кислорода и углекислого газа в крови, связывание их с гемоглобином и другими веществами и перенос с током крови.
4-й этап — диффузия газов в тканях (газообмен в тканях) — процессы, обеспечивающие диссоциацию оксигемоглобина в крови тканевых капилляров и диффузию кислорода из крови в тканевые структуры, а также диффузию углекислого газа в обратном направлении, его растворение и связывание с гемоглобином.
|
|
|
5-й этап — клеточное дыхание — биохимические и физико-химические процессы, обеспечивающие аэробное окисление органических веществ с получением энергии, используемой для жизнедеятельности клетки. При этом образуются углекислый газ, вода и азотистые основания (при окислении белков).
Такое выделение этапов дыхания удобно для последовательного его изучения. В клинической практике часто применяют иное подразделение, в частности под термином «внешнее дыхание» подразумевают как вентиляцию легких, так и газообмен между кровью и альвеолярным воздухом. В курсе физиологии человека изучаются преимущественно первые четыре этапа дыхания. Клеточное дыхание подробно изучается в курсе биохимии.
Внешнее дыхание
Вентиляция легких — процесс обмена воздуха между внешней средой и альвеолами легких.
Вентиляция легких (смена воздуха) осуществляется в результате периодических изменений объема грудной полости. Увеличение объема грудной полости обеспечивает вдох (инспирацию), уменьшение — выдох (экспирацию). Фазы вдоха и следующего за ним выдоха составляют дыхательный цикл.
Изменение объема грудной полости совершается за счет сокращений дыхательных мышц. Мышцы, при сокращении которых объем грудной полости увеличивается, называются инспираторными. К ним относятся диафрагма и наружные межреберные мышцы. При спокойном дыхании объем грудной клетки изменяется в основном за счет сокращения диафрагмы и перемещения ее купола. При глубоком форсированном дыхании в инспирации участвуют вспомогательные мышцы вдоха: трапециевидная, передние лестничные и грудино-ключично-сосцевидные мышцы. Спокойный выдох осуществляется в результате расслабления инспираторных мышц, что приводит к уменьшению объема грудной полости благодаря опусканию ребер (под действием силы тяжести) и расслаблению диафрагмы. Глубокий выдох происходит при сокращении экспираторных мышц, которыми являются внутренние межреберные мышцы и мышцы живота. К вспомогательным экспираторным мышцам относятся мышцы, сгибающие позвоночник.
|
|
|
Легкие располагаются в герметически замкнутой плевральной полости, которая образована наружным и внутренним листками плевры.
При спокойном дыхании давление в плевральной полости равно минус 6-8 мм рт. ст., т.е. на 6-8 мм рт. ст. ниже атмосферного. Его можно измерить, если ввести в щель иглу шприца, соединенного с манометром. Это обусловлено тем, что внутрилегочное давление равно атмосферному, а снаружи давление отсутствует или равно нулю. Эластическая сила легких уменьшает давление легких на пристеночную плевру. Следовательно, внутриплевральное давление равно
|
|
|
Рпл = Рлег — Рэл
где Рпл — внутриплевральное давление; Рлег — внутрилегочное давление, которое в состоянии покоя равно атмосферному; Рэл — эластическая сила легких.
В момент вдоха, когда сокращаются наружные межреберные мышцы и ребра поднимаются, наружный листок плевры отходит от внутреннего, вследствие чего увеличивается объем плевральной полости. Поскольку легкие всегда стремятся занять максимально возможный объем в грудной полости в связи с разностью давления внутри и снаружи органа, при увеличении объема плевральной полости происходят растяжение легких и поступление в них воздуха. Это приводит к увеличению эластической тяги легких и, следовательно, уменьшению внутриплеврального давления. Чем глубже вдох, тем больше уменьшается давление. В момент глубокого вдоха оно может достигать минус 12-15 мм рт. ст. (рис. 1).
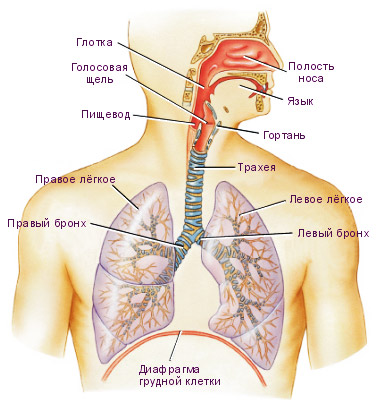
Когда в межреберных мышцах заканчивается процесс возбуждения, они расслабляются и ребра пассивно возвращаются в исходное положение; точно так же прекращение сокращения диафрагмы приводит к тому, что она занимает свое прежнее куполообразное положение. Возвращение ребер и диафрагмы в исходное положение приводит к уменьшению объема грудной полости, а следовательно, к сдавлению легких. При возвращении ребер в исходное положение давление в плевральной полости повышается, т.е. в ней уменьшается отрицательное давление, так как уменьшается эластическая тяга легких. При глубоком выдохе оно становится равным минус 3-4 мм рт. ст. При сдавлении легких из них пассивно выходит воздух — осуществляется выдох.
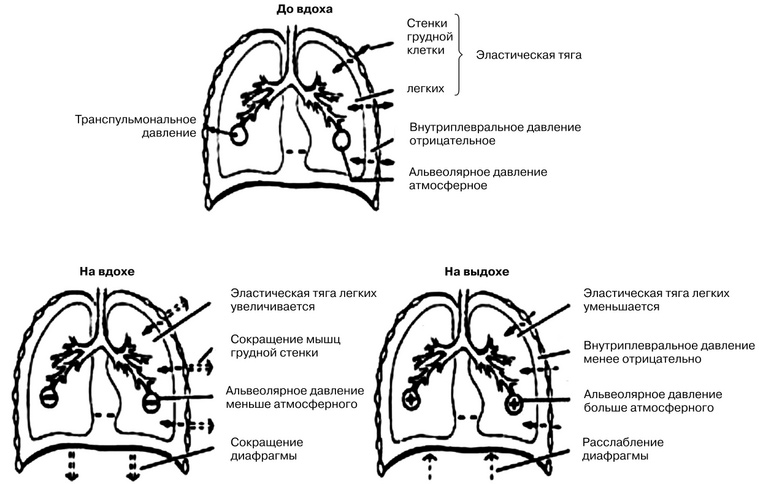
Рис. 1. Направления сил в течение дыхательного цикла
Упругие свойства легких. Эластическая тяга легких обусловлена тремя факторами:
§ поверхностным натяжением пленки жидкости, покрывающей внутреннюю поверхность альвеол;
§ упругостью ткани стенок альвеол вследствие наличия в них эластических волокон;
§ тонусом бронхиальных мышц.
Устранение сил поверхностного натяжения (заполнение легких солевым раствором) снижает эластическую тягу легких на 2/3
Если бы внутренняя поверхность альвеол была покрыта водным раствором, поверхностное натяжение должно было быть в 5-8 раз больше. В таких условиях наблюдалось бы полное спадение одних альвеол (ателектаз) при перерастяжении других. Этого не происходит потому, что внутренняя поверхность альвеол выстлана веществом, имеющим низкое поверхностное натяжение, так называемым сурфактантом, имеющим толщину 20-100 нм и состоящим из белков и липидов. Пленка сурфактанта обладает замечательным свойством: уменьшение размеров альвеол сопровождается снижением поверхностного натяжения; это важно для стабилизации альвеол.
Сурфактант необходим для начала дыхания при рождении ребенка. До рождения легкие находятся в спавшемся состоянии. Ребенок после рождения делает несколько сильных дыхательных движений, легкие расправляются, а сурфактант удерживает их от спадения (коллапса). Недостаток или дефекты сурфактанта вызывают тяжелое заболевание (синдром дыхательного дистресса). Поверхностное натяжение в легких у таких детей высокое, поэтому многие альвеолы находятся в спавшемся состоянии.
Показатели системы дыхания
Деятельность системы дыхания характеризуют определенные внешние показатели: частота дыхательных движений и легочные объемы.
Дыхательным объемом называют объем воздуха, который поступает в легкие при вдохе в состоянии покоя. Дыхательный объем у овец составляет 0,3-0,5 л, у лошадей — 4-6 л. Сверх данного количества животные могут вдохнуть еще определенный объем воздуха, который называется резервным объемом вдоха. После нормального выдоха животные могут выдохнуть приблизительно такое же количество воздуха. Этот объем называется резервным объемом выдоха. Объем воздуха, оставшийся в легких после выдоха резервного объема, называется остаточным объемом. Соответственно этому емкость легких называется общей.
Количество воздуха, которое животное или человек может максимально выдохнуть после самого глубокого вдоха, называется жизненной емкостью легких. Она складывается из дыхательного объема, резервного объема вдоха и резервного объема выдоха. Жизненная емкость у лошадей составляет до 26 л, у крупного рогатого скота — 30 л.
Жизненную емкость легких и дыхательные объемы можно определить с помощью спирометрии.
Объем воздуха, который остается в легких после спокойного выдоха, называется функциональной остаточной емкостью, или альвеолярным воздухом. Та часть общей емкости, которая вмещает дыхательный объем, называется емкостью вдоха.
Физиологическое мертвое пространство — воздух, который находится в воздухоносных путях (полости носа, носоглотке, трахее) и не участвует в газообмене. Хотя в воздухоносных путях не происходит газообмена, они необходимы для нормального дыхания, так как в них вдыхаемый воздух увлажняется, согревается, очищается от пыли и микроорганизмов. При раздражении пылевыми частицами и накопившейся слизью рецепторов носоглотки, гортани и трахеи возникает кашель, а при раздражении рецепторов полости носа — чиханье. Кашель и чиханье являются защитными дыхательными рефлексами.
Частота дыхательных движений или вентиляция легких определяется объемом воздуха, вдыхаемого или выдыхаемого в единицу времени. Количественной характеристикой легочной вентиляции является минутный объем дыхания — объем воздуха, проходящего через легкие за 1 мин.
Частота дыхательных движений в 1 мин в покое у лошадей составляет 8- 16, крупного рогатого скота — 10-30, свиней — 8-18. Минутный объем у лошадей достигает 40-60 л, у крупного рогатого скота — 25-30 л.
При адаптации организма к условиям внешней среды число дыхательных движений может увеличиться в 4-5 раз, дыхательный объем воздуха — в 4-8 раз, минутный объем дыхания — в 10-25 раз.
Газообмен в легких
Газообмен в легких между альвеолярным воздухом и кровью капилляров малого круга кровообращения осуществляется в результате диффузии O2 из альвеолярного воздуха в кровь и СО2, из крови в альвеолярный воздух. Диффузия происходит вследствие разности парциального давления (давление отдельного газа в смеси газов) этих газов в альвеолярном воздухе и в крови. Парциальное давление O2 в альвеолярном воздухе составляет около 100 мм рт. ст. Напряжение O2 в венозной крови равно 40 мм рт. ст. В результате этой разницы парциального давления O2 из альвеолярного воздуха поступает в кровь. Напряжение СО2, в венозной крови, поступающей к легким, составляет 46 мм рт. ст., а парциальное давление СO2 в альвеолярном воздухе — около 40 мм рт. ст. Вследствие этого СO2 поступает из крови в альвеолярный воздух до выравнивания его парциального давления в крови и в альвеолярном воздухе.
Газообмену в легких способствует большая поверхность альвеол и малая толщина легочной мембраны. В течение суток из альвеол в кровь переходит у коровы около 5000 л О2, а из крови в альвеолярный воздух поступает около 4300 л СО2.
Контакт крови с альвеолярным воздухом происходит за 0,3-0,7 с, и за этот период парциальное давление газа полностью выравнивается.
Транспорт газов кровью
В нормальных условиях транспорт кислорода и диоксида углерода кровью осуществляется в растворенном и химически связанном виде. Из общего количества кислорода, который содержится в артериальной крови, только 0,3% его растворено в плазме, а остальное количество находится в химической связи с гемоглобином. Поэтому главное значение имеет транспорт дыхательных газов в связанном с гемоглобином состоянии.
Транспорт кислорода кровью осуществляется в основном за счет обратимого присоединения молекул O2 к молекуле гемоглобина. Более 90% кислорода крови, выходящей из легких, переносится в виде оксигемоглобина (НЬO2). Одна молекула гемоглобина присоединяет к себе четыре молекулы кислорода; 1 г гемоглобина способен присоединить 1,34 мл кислорода. Максимальное количество кислорода, которое может связать кровь при полном насыщении гемоглобина кислородом, называется кислородной емкостью крови. В нормальных условиях количество кислорода, связанного с гемоглобином, зависит от парциального напряжения кислорода в крови или тканях. При уменьшении парциального напряжения кислорода в среде количество гемоглобина, находящегося в форме оксигемоглобина, уменьшается. Гемоглобин представляет собой белок, состоящий из четырех субъединиц, каждая из которых содержит один гем. Гем — протопорфин, состоящий из четырех пиррольных колец, связанных между собой метиловыми мостиками. В центре гема находится двухвалентное железо. Одна молекула гемоглобина присоединяет к себе четыре молекулы кислорода.
Сродство гемоглобина к кислороду возрастает при высоком парциальном давлении последнего. Такие условия создаются в легких, где почти весь гемоглобин (98%) насыщается кислородом. Сродство гемоглобина к кислороду снижается при увеличении концентрации СО2 и Н+. Такие условия создаются в тканях, где интенсивно протекают процессы обмена веществ. Оксигемоглобин поэтому быстро диссоциирует, O2 освобождается и поступает в ткани, где его напряжение значительно ниже, чем в артериальной крови (100 мм рт. ст.). В венозной крови напряжение O2 составляет всего 35-45 мм рт. ст. Протекая по тканевым капиллярам, кровь отдает кислород тканям.
Графическое изображение степени насыщения гемоглобина кислородом, т.е. образование оксигемоглобина, называется кривой диссоциации оксигемоглобина. Она имеет S-образную форму. Кривая диссоциации гемоглобина может сдвигаться вправо при повышении парциального давления СО2 и снижении рН. Это уменьшает сродство гемоглобина к кислороду и улучшает его отдачу в тканях. Сдвиг влево происходит при снижении парциального давления СO2 и повышении рН. В этом случае увеличивается сродство гемоглобина к кислороду и ухудшается его отдача в тканях.
Диоксид углерода в крови транспортируется в трех видах: физически растворенном (2-3%), химически связанном в виде бикарбонатов (80%) и химически связанном с гемоглобином в виде карбгемоглобина (4-5%).
Когда капиллярная кровь протекает по тканям с высоким парциальным давлением диоксида углерода, последний устремляется в кровь и растворяется в плазме. Диоксид углерода быстро диффундирует из плазмы крови в эритроциты. Соединяясь с водой, он образует слабую угольную кислоту. В плазме крови эта реакция протекает замедленно, а в эритроцитах под влиянием фермента карбоангидразы реакция ускоряется в несколько тысяч раз. Здесь угольная кислота быстро диссоциирует на ионы Н+ и НСО3-, и большая часть НСО3-, снова выходит в плазму крови. Основная масса свободных ионов водорода связывается с дезоксигемоглобином. Одновременно дезок- сигемоглобин теряет сродство к ионам калия, поэтому эти ионы освобождаются и идут на образование КНСО3,. Бикарбонат свободно диффундирует через мембрану эритроцита в окружающую плазму в силу разницы концентраций этого аниона. Благодаря избирательной проницаемости мембраны эритроцита диффузия бикарбоната создает трансмембранную разность потенциалов. С учетом того, что в эритроците образуется большое количество анионов НСО3-, часть их выходит из эритроцитов в плазму крови, где связывается с ионами натрия, образуя бикарбонат натрия. В обмен на вышедшие анионы НСО3-, внутрь эритроцитов проникают анионы хлора. Поэтому эритроциты наывают фабрикой бикарбонатов. В целом же, пройдя через эритроцит, угольная кислота в итоге превращается в бикарбонат натрия в плазме крови и бикарбонат калия в эритроцитах и в таком виде переносится к легким. Одновременно в эритроците небольшая часть СO2 образует карбаминовую связь с гемоглобином и в результате переносится внутри эритроцитов в виде карбгемоглобина. В целом в капиллярах легких при низком парциальном давлении и напряжении диоксида углерода происходит процесс, направленный на выделение присоединенного в тканях С02. Напряжение С02, в клетках может достигать 60 мм рт. ст. В тканевой жидкости оно в среднем составляет 46 мм рт. ст. Диффундируя в направлении более низкого напряжения, СO2 переходит из клеток в тканевую жидкость, а далее в кровь и делает ее венозной.
Дыхание в условиях пониженного и повышенного атмосферного давления
Дыхание при пониженном атмосферном давлении. При подъеме на высоту животные и человек оказываются в условиях пониженного атмосферного давления. При этом развивается гипоксия (недостаток кислорода в организме) в результате низкого парциального давления кислорода во вдыхаемом воздухе. На высоте 5 км барометрическое давление составляет около 60 мм рт. ст. и насыщенность крови кислородом снижается до 80% , что способствует развитию горной болезни.
На высоте от 2,5 до 5 км повышается вентиляция легких, что вызвано стимуляцией каротидных хеморецепторов. Одновременно происходит повышение артериального давления и увеличение частоты сердечных сокращений. Эти реакции направлены на усиление снабжения тканей кислородом.
В случае увеличения высоты более 7 км могут наступить опасные для жизни нарушения дыхания, кровообращения и потеря сознания.
Длительное пребывание или обитание животных и людей в горной местности сопровождается акклиматизацией к кислородному голоданию, которая проявляется в следующем:
§ увеличивается концентрация эритроцитов в крови в результате усиления эритропоэза;
§ повышается содержание гемоглобина в крови и увеличивается ее кислородная емкость;
§ активизируется вентиляция легких;
§ повышается плотность кровеносных капилляров в тканях в результате увеличения их длины и извитости.
Дыхание при повышенном атмосферном давлении. При погружении животных и человека под воду возрастает атмосферное давление. Например, на глубине 10 м давление возрастает до 2 атм, на глубине 20 м — до 3 атм. В этом случае парциальное давление газов в альвеолярном воздухе возрастает и в крови растворяется большое количество газов — кислорода, азота. Само пребывание на большой глубине не опасно, но при быстром подъеме и переходе от повышенного давления к обычному растворенные в крови газы вскипают и вызывают газовую эмболию сосудов (кессонная болезнь), что может привести к смерти. Кессонная болезнь характеризуется болями в мышцах, головокружением, одышкой, потерей сознания. При медленном подъеме на поверхность газы постепенно удаляются из организма, что профилактирует развитие кессонной болезни. Особенно важны эти закономерности при проведении водолазных работ. В случае погружения водолазов на большие глубины для дыхания применяют гелиево-кислородные смеси. Водолазы поднимаются с глубины очень медленно, а после подъема проходят постепенную декомпрессию.
У некоторых животные выработались специальные дыхательные приспособительные реакции, позволяющие им нырять на определенную глубину. К таким животным относятся ластоногие, киты, выдра, калан и многие другие. Например, крупные киты могут погружаться на глубину 100-200 м и находиться под водой в течение 50-60 мин, а морские львы могут нырять на глубину до 750 м. Физиологически это обусловлено тем, что их дыхательный центр малочувствителен к накоплению в организме СО2, что позволяет длительно задерживать дыхание и более полно использовать O2, содержащийся в крови и легких. Кроме того, их мышцы богаты миоглобином. Миоглобин — красный железосодержащий белок (специализированная разновидность гемоглобина), находящийся в сердечной и скелетной мышцах и активно переносящий O2. Так, в скелетных мышцах лошадей и человека содержится 4-9 мг миоглобина на 1 г массы мышц, а у морских львов — 55-75 мг/г.
Дата добавления: 2020-11-29; просмотров: 169; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!