Роль каналов пространственной частоты
В функционировании зрительной системы человека
Рассмотрим рис. 4. Левая верхняя решетка (рис. 4, А) имеет сравнительно низкую пространственную частоту (широкие полосы), а нижняя — высокую (узкие полосы). Пространственные частоты (ширина полос) решеток на рис. 4, В идеи-
Шиффман X . Анализ пространственной частоты
103
 Рис. 4. Паттерны, рассматриваемые испытуемым, А и тестовые паттерны Б, используемые для доказательства наличия в зрительной системе специализированных каналов, предназначенных для восприятия пространственной частоты. Методика проведения эксперимента описана в тексте1
Рис. 4. Паттерны, рассматриваемые испытуемым, А и тестовые паттерны Б, используемые для доказательства наличия в зрительной системе специализированных каналов, предназначенных для восприятия пространственной частоты. Методика проведения эксперимента описана в тексте1
тичны и занимают промежуточное положение между пространственными частотами решеток, представленных на рис. 4, А. Прикройте решетки на В и не менее 60 с внимательно рассматривайте решетки на А, переводя взгляд слева направо и обратно вдоль горизонтальной линии фиксации, разделяющей два паттерна. После завершения периода адаптации переведите взгляд на точку фиксации между двумя решетками (средняя пространственная частота) на Б. Пространственные частоты теперь уже не будут казаться идентичными: пространственная частота верхнего паттерна будет казаться выше пространственной частоты нижнего.
Иными словами, снижение чувствительности к определенной пространственной частоте (или нескольким определенным пространственным частотам) вследствие их избирательного воздействия подтверждает наличие в зрительной системе отдельных каналов, воспринимающих разные пространственные частоты.
|
|
|
Большинство объектов, воспринимаемых зрительной системой, может быть проанализировано с точки зрения их пространственной частоты. Описанный ниже анализ пространственной частоты сложных форм — еще одно подтверждение роли анализа Фурье в обработке зрительной информации.
1 См.: Blakemore С , Sutton P. Size adaptation: A New Aftereffect // Science. 1969. 166. P. 245.
104
Тема 17. Экспериментальные исследования восприятия
Обработка зрительной информации: блок-портреты
Пример, иллюстрирующий роль анализа пространственной частоты в восприятии формы, представлен на рис. 5, А.
Фотография, так называемый блок-портрет, президента США представляет собой результат специальной компьютерной обработки традиционного портрета. Исходный портрет после обработки на компьютере был поделен на маленькие квадраты, или блоки, причем каждый из этих блоков был так воспроизведен типографским способом, что соответствующие участки исходного портрета сохранили присущую им среднюю интенсивность света и тени. Иными словами, светимость каждого блока одинакова (однородна) на всей его площади, т.е. она была поэлементно усреднена, или оцифрована1. Таким образом исходный портрет был превращен в набор примыкающих друг к другу светлых и темных блоков. При этом портрет лишился многих деталей и первоначальной четкости, поскольку вследствие поэлементного усреднения светимости всех блоков
|
|
|

Рис. 5. А — блок-портрет, полученный на компьютере; В — тот же самый портрет после удаления высокочастотного шума
1 См.: Harmon L.D. The recognition of faces // Scientific American. 1973. 229. P. 70-82; Harmon L.D., Julesz B. Masking in visual recognition: Effects of two-dimensional filtered noise // Science. 1973. 180. P. 1194-1197.
Шиффман X . Анализ пространственной частоты
105
оригинала были удалены высокие пространственные частоты. Одновременно с этим резкие границы между блоками стали причиной возникновения своего рода высокочастотного шума — нежелательного последствия процесса усреднения светимостей блоков. Этот шум тоже маскирует многие информативные низкочастотные компоненты, оставшиеся от оригинального портрета.
В результате при прямом и пристальном взгляде на портрет на нем не сразу разглядишь легкоузнаваемое лицо. Узнавание значительно облегчается, если портрет оказывается не в фокусе, т.е. если смотреть на него с некоторого расстояния или сбоку. Очевидное объяснение этого явления заключается в том, что при искусственной «расфокусировке» блок-портрета избирательно отфильтровывается большая часть высокочастотного шума, образовавшегося в процессе создания блок-портрета, и общий уровень шума понижается. В результате появляется возможность воспринимать многие из оставшихся низких частот. Иными словами, при тех условиях, при которых портрет оказывается не в фокусе, уменьшается видимость искусственных резких границ между однородными квадратами, или блоками, которые образовались в результате оцифровки последних. Устранение привнесенных оцифровкой высокочастотных деталей делает видимыми низкочастотные информативные детали, что и облегчает узнавание лица. Графически это представлено на рис. 5, Б.
|
|
|
Пространственная частота и острота зрения
Анализ функционирования зрительной системы, учитывающий ее чувствительность к светлотному контрасту, является основой для выработки более информативного и полного критерия оценки остроты зрения, например остроты разрешающей способности глаза, <...> оцененной исключительно при одной пространственной частоте и при одном уровне светового контраста. Например, у человека может быть средняя чувствительность к одним пространственным частотам и чувствительность ниже средней — к другим. Известно, что возраст по-разному влияет на контрастную чувствительность к разным пространственным частотам: чувствительность к высоким частотам значительно снижается, а чувствительность к низким остается практически неизменной1. Следовательно, в результате старения утрачивается способность различать мелкие детали.
|
|
|
1 См.: Owsley С, Sekuler R ., Slemsen D . Contrast sensitivity throughput adulthood // Vision Research. 1983. 23. P. 689-699; Crassini В ., Brown В ., Bowman K. Age-related changes in contrast sensitivity in central and peripheral retina // Perception. 1988. 17. P. 315-332.
106
Тема 17. Экспериментальные исследования восприятия
Аналогично возрасту действует на контрастную чувствительность к высоким пространственным частотам и движение — оно ее понижает. Иными словами, при стимулировании движущимся раздражителем, образованным высокими пространственными "частотами, восприятие мелких деталей ухудшается1. Обсудим практические последствия этого явления для такого вида деятельности, как управление транспортным средством. Хотя способность видеть другие легковые автомобили и грузовики (низкие пространственные частоты) при движении в потоке машин остается неизменной, способность различать надписи на дорожных указателях, а также растущие вдоль дороги деревья и кусты (высокие пространственные частоты) понижается. Как правило, подобные изменения остроты зрения не фиксируются ни с помощью <...> таблицы Снеллена, <...> ни с помощью таблицы, которая помогла установить, что у детей способность к распознаванию большинства пространственных частот выражена слабее, чем у взрослых2.
Исследования, выполненные Гинсбургом, показывают, что острота зрения людей, чья профессия предъявляет к ней повышенные требования, например острота зрения летчиков, измеренная традиционными методами, вполне может оказаться нормальной или средней, однако если оценить ее, учитывая и контрастную чувствительность, обнаруживаются весьма существенные индивидуальные различия3. Автор нашел, что некоторые летчики демонстрируют большую контрастную чувствительность в случае низких пространственных частот, чем их коллеги, острота зрения которых по итогам теста Снеллена, в ходе которого предъявляется высокочастотный стимул ьный материал, была признана более высокой4. Подобная ярко выраженная избирательная чувствительность может проявляться в улучшении восприятия объектов при плохой видимости, например в тумане, или объектов, находящихся на большом рас-
1 См.: Long G.M., Homolka J.L. Contrast sensitivity during horizontal visual pursuit:
dynamic sensitivity functions // Perception. 1992. 21. P. 753-764; Long G.M., Kearns D.F.
Visibility of text and icon highway signs under dynamic viewing conditions // Human Factors.
1996. 38. P. 690-701.
2 См.: Dobson V., Teller D.Y. Visual acuity in human infants: A review and comparison of
behavioral and electrpohysical studies // Vision Research 1978. 18. P. 1469-1483; GwiazdaJ.,
Brill S., Held R. New metods for testing infant vision // The Sigbtsaving Review. 1979. 49.
P. 61-69; Norcia A.M., Tyier C.W, Spatial frequency sweep VEP: Visual acuity during the
first year of life // Vision Eesearch. 1985. 25. P. 1399-1408.
3 См.: Oinsburg A.P. Spatial filtering and vision: Implications for normal and abnormal vision // Ь.Ргоеша, J.Enoch, A.Jampolski (Eds.). Clinical applications of psychophysics. N. Y.: Cambridge University Press, 1981; Ginsburg A.P. Spatial filtering and visual form perception // K.R.Boff, L.Kaufman, J.P.Thomas (Eds.). Handbook of perception and human performance. Vol. II: Cognitive processes and performance. N. Y.: John Wiley, 1986.
4 См.: Ginsburg A.P, Spatial filtering and vision: Implications for normal and abnormal vision // L. Proenza, J. Enoch, A. Jampolski (Eds.). Clinical applications of psychophysics. N. Y.: Cambridge University Press, 1981.
Шиффман X. Анализ пространственной частоты
107
стоянии. На основании результатов тестирования по Снеллену Гинсбург предположил, что от 10 до 15% населения имеет хорошую остроту зрения, но плохую чувствительность к низким и средним пространственным
частотам1.
Роль анализа пространственной частоты
в зрительном восприятии
Какую роль в изучении зрительного восприятия играет анализ пространственной частоты? Прежде всего он является простым и надежным способом описания и обобщения структурных деталей различных визуальных объектов, а именно: высокие пространственные частоты кодируют информацию о деталях, обладающих наиболее тонкой текстурой, а низкие пространственные частоты — информацию о структурах, образованных паттернами крупных элементов. С его помощью можно также описать и общие принципы работы визуальной системы, связанной с анализом, сопоставлением и интеграцией активности огромного количества рецепторов и соотнесением этой активности со специфическим признаком физического раздражителя. Кроме того, мы убедились, что описание визуального раздражителя в терминах его пространственных частот полезно для оценки остроты зрения и более информативно, чем оценка последней с помощью традиционной таблицы Снеллена. Анализ пространственной частоты не просто выявляет, какие именно количественные и дескриптивные характеристики может использовать зрительная система для кодирования сложной визуальной информации, подлежащей дальнейшей обработке, но играет в зрении более существенную роль.
1 См.: GimburgA.P. Spatial filtering and visual form perception // K.R.Boff, L.Kaufman, J.P.Thomas (Eds.). Handbook of perception and human performance. Vol. II: Cognitive processes and performance. N. Y.: John Wiley, 1986; Ginsburg A.P., Cannon M.W., Evans D.W., Owsley C, Mulvaney P. Large sample norms for contrast sesitivity // American Journal of Optometry and Physiological Optics. 1984. 61. P. 80-84.
А . Д . Логвиненко
ПЕРЕДАТОЧНАЯ ФУНКЦИЯ ВСЕЙ ЗРИТЕЛЬНОЙ СИСТЕМЫ В ЦЕЛОМ1
Основное внимание <...> мы уделим анализу методов и результатов эмпирического построения полной передаточной функции, поскольку это составляет центральную задачу данной главы.
Многообразие методов идентификации линейных систем2 обусловило и многообразие психофизических методов идентификации полной передаточной функции. В силу ряда причин в психофизике зрения наибольшее распространение получили частотные методы, в частности, один из вариантов метода гармонического стимула, заключающегося с точки зрения психолога в построении изоконтрастных кривых. Это связано с тем, что для идентификации линейной и инвариантной системы методом гармонического стимула, вообще говоря, не обязательно иметь функцию светлоты тестового гармонического стимула. <...> Для такой системы образ гармонического стимула также будет гармоническим и для идентификации системы достаточно знать его амплитуду и фазу. Принимая во внимание необычайную трудоемкость процедуры построения функции светлоты для трехмерного стимула l {х', х2, t ), легко понять не только привлекательность метода гармонического стимула для психологов, но и то, почему идентификации подвергалась не вся трехмерная передаточная функция зрительной системы в целом: H ( f , f , w ), а лишь ее одномерные или двумерные сечения.
Первые попытки эмпирического построения передаточной функции зрительной системы человека были предприняты более полувека назад (кстати сказать, задолго до появления математической теории идентификации систем как таковой). С помощью порогового варианта метода
1 Фурье-анализ зрительного восприятия / Под ред. А.Д.Логвиненко. М.: Изд-во Моск. ун-та, 1982. С. 33-38.
- См.: Эйкхофф П. Основы идентификации систем управления. М., 1975.
Логвиненко А , Д . Передаточная функция всей зрительной системы в целом 109
изоконтрастных кривых определялась временная передаточная функция Н (0,0, w)1. Четверть века назад этим же методом была построена одномерная пространственная — Н ( f1, 0, 0) передаточная функция2. Пространственно-временную — Н ( f1, 0, w) передаточную функцию впервые построил также этим методом Дональд Келли3.
Метод изоконтрастных кривых
Простоты ради и следуя сложившейся традиции изложение этого метода будем вести для одномерного случая, например, для идентификации пространственной передаточной функции Н ( f, 0, 0). Не изменяющиеся аргументы в дальнейшем будем, как правило, опускать, т.е. писать H( f) вместо H( f, 0, 0) или l(х) вместо l( x') l( x2) l( i) u l( x, t) вместо l (х1, t) 1 (хх).
Важной характеристикой любого метода идентификации является выбор тестовых стимулов. Для метода изоконтрастных кривых такими стимулами являются вертикальные синусоидальные решетки, яркостные функции которых имеют следующий вид (рис. 1):

Синусоидальная решетка (1) вполне определяется тремя числами: ее средней яркостью ( lо), контрастом (т) и пространственной частотой ( f). Контраст решетки, так называемый контраст Майкельсона, связан с наиболее ( lтах) и наименее ( lmin) яркими участками решетки следующим соотношением:

Пространственная частота есть величина, обратная периоду D (рис.1) решетки, т.е. f = D~'. Поскольку для проксимального стимула пространственные координаты измеряются в угловых единицах (градусах или минутах), то размерность пространственной частоты, следовательно, угл. град.-1 или угл. мин.-1. (Отметим попутно, что временные частоты измеряются в герцах — Гц). <...>
1 См.: Ives H . E . A theory of intermittent vision // J. Opt. Soo. Am. and Rev. Sci. Instr.
1922. 6. P.343-361; De Lange. Experiments on flicker and some calculations on an electrical
analogue of the foveal systems // Physics. 1952. 18. P. 935-950.
2 См.: Schade O.H. Optical and photoelectric analog of the eye //' J. Opt. Soc. Am. 1956.
46. P. 721-739.
3 См.: Kelly D.H. Frequency doubling in visual responses // J. Opt. Soc. Am. 1966. 56.
P. 1628-1633.
| 110 Тема 17. Экспериментальные исследования восприятия |

Рис. 1. Яркостная функция для синусоидальной решетки
В основе метода изоконтрастных кривых лежат следующие рассуждения. Передаточная функция зрительной системы показывает, насколько система изменяет контраст синусоидальной решетки. Поэтому ее определение сводится к вычислению отношения субъективного контраста решетки к контрасту физической решетки как функции пространственной частоты. При этом можно поступать двояким способом. Либо зафиксировать контраст физической решетки (т.е., предъявлять синусоидальные решетки с разной пространственной частотой, имеющие одинаковый контраст) и измерять субъективный контраст как функцию пространственной частоты; либо, задавшись некоторым определенным уровнем субъективного контраста, экспериментально определить для каждой пространственной частоты тот физический контраст, при котором решетка имеет заданную величину субъективного контраста. Полученная в результате кривая, показывающая, какую величину физического контраста на той или иной пространственной частоте необходимо взять для того, чтобы субъективный контраст был одним и тем же на любой пространственной частоте, называется изоконтрастной кривой. Изоконтрастная кривая есть функция, обратная к передаточной функции (с некоторым коэффициентом пропорциональности).
Непременным условием применимости метода изоконтрастных кривых для построения передаточной функции зрительной системы является следующее соотношение;
т ( f 3 ) – т0 ( f 3 )- m ( f )- m 0 ( f ), (3)
где т ( f 3 ) — фиксированный контраст эталонной решетки с постоянной пространственной частотой f 3 ; m ( f ) — контраст решетки с переменной пространственной частотой /, подобранный таким образом, что субъективные контрасты обеих решеток равны; m 0 ( f 3 ), m 0 ( f ) — пороговые контрасты эталонной и переменной решеток соответственно.
Это равенство означает, что любая изоконтрастная кривая должна получаться из пороговой кривой сдвигом вдоль оси ординат на величи-
Логвиненко А.Д . Передаточная функция всей зрительной системы в целом 111
ну, пропорциональную контрасту стандартной решетки. Именно такое соотношение между пороговыми и изоконтрастными кривыми было получено экспериментально1. Однако этот автор помимо эталонной решетки с частотой 5 угл. град, использовал всего лишь две переменные решетки 1,67 и 15 угл. град.-1. Ясно, что три точки слишком мало для того, чтобы судить о выполнении условия (3). Более того, ряд исследователей указывают на то, что форма изоконтрастнойкривой изменяется с изменением контраста эталонной решетки2. Так, по данным некоторых авторов, изоконтрастные кривые становятся практически горизонтальными линиями для эталонных решеток с контрастом, близким к единице3. Поэтому изоконтрастные кривые могут служить средством построения передаточной функции зрительной системы лишь при условии малости контраста эталонной решетки. Только в этом случае можно надеяться на выполнение равенства (3).
 Наименьший доступный испытуемому контраст эталонной решетки — это ее пороговый контраст. В некотором смысле кривую пороговых контрастов можно рассматривать как разновидность изоконтрастной кривой. Действительно, согласно существующим ныне представлениям о механизме обнаружения <...> испытуемый в пороговой ситуации отвечает реакцией «да», если субъективный контраст решетки превышает некоторый фиксированный уровень , называемый критерием испытуемого. Существуют некоторые косвенные свидетельства в пользу того, что критерий е не зависит от частоты решетки4. Если это так, то кривая пороговых контрастов — это изоконтрастная кривая, соответствующая субъективному контрасту, равному критерию испытуемого.
Наименьший доступный испытуемому контраст эталонной решетки — это ее пороговый контраст. В некотором смысле кривую пороговых контрастов можно рассматривать как разновидность изоконтрастной кривой. Действительно, согласно существующим ныне представлениям о механизме обнаружения <...> испытуемый в пороговой ситуации отвечает реакцией «да», если субъективный контраст решетки превышает некоторый фиксированный уровень , называемый критерием испытуемого. Существуют некоторые косвенные свидетельства в пользу того, что критерий е не зависит от частоты решетки4. Если это так, то кривая пороговых контрастов — это изоконтрастная кривая, соответствующая субъективному контрасту, равному критерию испытуемого.
Величина, обратная пороговому контрасту, называется контрастной чувствительностью. Зависимость контрастной чувствительности от пространственной частоты синусоидальной решетки будем называть функцией контрастной чувствительности. Таким образом, мы приходим к выводу, что функция контрастной чувствительности совпадает с передаточной функцией зрительной системы с точностью до некоторого постоянного множителя. В дальнейшем разновидность метода, связанного с построением функции контрастной чувствительности, будем называть пороговым методом идентификации передаточной функции.
1 См.: Kulikowshi J.J. Effective contrast constancy linearity of contrast sensation //
Vision Research. 1976. 16. P. 1419-1431.
2 См.: Watanabe A., Mori Т ., Nagata S., Hiwatashi K. Spatial sine-wave responses of the
human visual system // Vision Research. 1968. 8. P, 1245-1264; Blakemore C„ Campbell F.W,
On the existence of neurones in the human visual system selectively sesitive to the orientation
and size of retinal images /'/ J. Physiol. 1969. 205. P. 237-260.
3 См.: Georgeson MA., Sullivan G.D. Contrast constancy: deblurring in human vision by spatial frequence channels // J. Physiol. 1975. 252. P. 627-656.
4 См.: Sachs M., Nachmias J., Robson J.G. Spatial-frequency channels in human vision // J. Opt. Soc. Am. 1971. 61. P. 1176-1186.
112
Тема 17. Экспериментальные исследования восприятия
Метод изоконтрастных кривых для идентификации передаточной функции зрительной системы, как правило, применяют в его пороговом варианте. Одним из немногих авторов, которые использовали непороговый вариант этого метода, является О. Врингдал1. Он определял отношение контрастов (субъективного к физическому) при различных величинах физического контраста для пространственных частот в диапазоне 0,5—9,0 угл.град.-1 со средней яркостью 0,25:20 нит методом подравнивания. Испытуемого просили подравнять яркость однородного поля к максимальной яркости, которую имеют светлые полосы синусоидальной решетки. Затем испытуемый должен был установить яркость однородного поля таким образом, чтобы его светлота равнялась наиболее темному участку на синусоидальной решетке. Отношение этих величин принималось им за величину субъективного контраста. Построенные этим автором кривые, показывающие зависимость отношения субъективного к объективному контрасту от пространственной частоты примечательны в двух отношениях. Во-первых, все они имеют максимум на частоте 5—7 угл.град.-1. Во-вторых, отношение контрастов превышает единицу для всех пространственных частот исследовавшегося диапазона, т.е. зрительная система в этом диапазоне усиливает контраст. Явление усиления контраста хорошо известно в психологии зрительного восприятия2. Однако, как правило, это явление связывается с более сложными процессами обработки зрительной информации, нежели обсуждаемые здесь. Представляется необходимым подвергнуть явление усиления контраста синусоидальной решетки дополнительному исследованию с применением более тонких психофизических методов.
1 См.: Bryngdahl О . Characteristics of the visual system: psychophysical measurements
of the responce to spatial sine-wave stimuli in the mesopic region // J. Opt. Soc. Am. 1964.
54. P. 1152-1160; Bryngdahl O. Regular occurence of simultaneous brightness contrast in
the mesopic region // Kybernetik. 1965. 2. P. 227-236.
2 См.: Heineman E.G. Simultaneous brightness induction /7 D.Jameson, L.M.Hurvich
(Eds.). Handbook of Sensory Physiology. N. Y., 1972. Vol. VII/4. Visual Psychophysics.
Часть 2. Факты, закономерности и результаты
Исследований восприятия
1. Восприятие цвета. Основные субъективные характеристики цвета и их объективные корреляты. Адаптация. Явление Пуркинье. Виды контрастов. Законы смешения цветов. Цветовой круг и треугольник. Цветовое тело. Теории цветового зрения
Б . М . Величковский ,
В. П. Зинченко,
А . Р . Лурия
ВОСПРИЯТИЕ ЦВЕТА1
Зрительная система человека чувствительна к электромагнитным колебаниям, длина волны которых лежит в диапазоне от 380 до 720 нм (миллионных долей миллиметра). Эта область электромагнитных колебаний называется видимой частью спектра.
Рецепция падающего на сетчатку света представляет собой только первую ступень в сложной цепи процессов, приводящих к зрительному отражению окружающего нас мира. Структура процесса восприятия цвета меняется в зависимости от оптических свойств поверхностей предметов, которые должны быть восприняты наблюдателем. Эти поверхности могут светиться, излучая больше света, чем на них падает; блестеть, отражая весь падающий на них свет; отражать лишь часть падающего света и, наконец, быть прозрачными, т.е. не оказывать свету существенных препятствий.
Значительное большинство окружающих нас предметов относится к группе тел, частично поглощающих и частично отражающих падающий на них от искусственных или естественных источников свет. Цвет этих предметов объективно характеризуется их отражательной способностью. Поэтому для восприятия цвета предметов зрительная система должна учитывать не только свет, отраженный поверхностью предмета, но также характеристики освещающего эту поверхность света.
Однако в том случае, когда поверхность светится или специально исключены признаки ее принадлежности какому-либо предмету, восприятие цвета может основываться лишь на анализе непосредственно излучаемого поверхностью света. Эта ситуация имеет место, если участок
1 Величковский Б.М., Зинченко В.П., ЛурияА.Р. Психология восприятия. М.: Изд-во Моск. ун-та, 1973. С. 81, 83, 109-126.
114
Тема 17. Экспериментальные исследования восприятия
поверхности рассматривается через отверстие в большом темном или сером экране. Благодаря экрану скрадывается расстояние до поверхности и воспринимается диффузный цвет, относительно равномерно заполняющий отверстие. Такие цвета называются апертурными. Благодаря относительной простоте процессов восприятия апертурных цветов, они изучены в настоящее время более полно, чем восприятие цвета предметов. Кроме того, исследование восприятия апертурных цветов имеет важное практическое значение, так как именно с ними приходится иметь дело операторам, работающим с современными индикаторами. Широкое использование цвета для кодирования информации связано с относительной легкостью различения апертурных цветов.
Цветовые ощущения, возникающие при восприятии апертурных цветов, полностью описываются тремя характеристиками или качествами. К ним относятся светлота, цветовой тон и насыщенность.
Первая из этих характеристик — светлота — иногда также называется видилюй яркостью. Светлота определяется прежде всего физической яркостью света. Как показали психофизические исследования, зрительная система способна реагировать на очень незначительные изменения яркости света: дифференциальный порог яркости равен всего лишь 0,01.
Измеряемые с помощью психофизических методов количественные отношения между интенсивностью раздражителя и величиной ощущения не остаются постоянными. В зависимости от условий, в которых осуществляется восприятие, происходит изменение как абсолютной, так и разностной чувствительности.
Важнейшим фактором, определяющим уровень чувствительности, является интенсивность действующих на организм раздражителей. Например, изменение освещенности предметов в течение суток настолько значительно, что будь чувствительность глаза неизменной, человек либо оказывался слепым на ярком солнечном свете, либо был совершенно неспособен к восприятию в сумерках. Этого не происходит потому, что в условиях недостаточной освещенности абсолютная зрительная чувствительность обостряется, а на ярком свету — снижается. Подобное приспособительное изменение чувствительности в зависимости от условий среды называется адаптацией.
<...>
Анализ динамики световой чувствительности при адаптации к темноте позволяет установить момент перехода от колбочкового к палочковому зрению. Для этого адаптированного к дневному свету наблюдателя помещают в полную темноту и периодически измеряют нижний абсолютный порог яркости. Результаты измерений показывают, что вначале порог быстро падает, стабилизируясь на постоянном уровне через 8— 10 мин, а затем наступает вторичное резкое снижение порога, которое прекращается лишь через 30—40 мин после начала адаптации (рис. 1).
Величковский Б . М ., Зинченко В . П ., Лурия А . Р . Восприятие цвета 115

Рис. 1. Изменение нижнего абсолютного порога яркости
в ходе зрительной темновой адаптации:
I — тестирование белым светом; II — тестирование красным светом
Такой «двухступенчатый» вид кривая темновой адаптации имеет только тогда, когда пороги тестируются белым светом. Если используется красный свет, к которому палочки нечувствительны, кривая адаптации состоит только из своей первой ветви. Это доказывает, что точка перелома на кривой темновой адаптации соответствует моменту перехода от колбочкового к палочковому зрению.
Процесс световой адаптации продолжается обычно всего лишь доли секунды.
Видимая яркость меняется также в зависимости от длины волны раздражителя.
При дневном освещении более яркими кажутся тона, сдвинутые к длинноволновой, красной части спектра. В сумерках же кривая спектральной чувствительности сдвигается в сторону коротковолнового конца видимого спектра (рис. 2). В этом случае наблюдается потемнение крас-
Д

Рис. 2. Кривые спектральной чувствительности глаза в темноте (а) и на свету (б).
116
Тема 17, Экспериментальные исследования восприятия
|
|

|
|

Рис. 3. Яркостный контраст
ного и высветление синего тонов. Это явление носит имя открывшего его чешского физиолога Яна Пуркинье.
Большой интерес представляют явления зрительного контраста. Одновременный или пространственный яркостный контраст заключается в подчеркивании зрительной системой различий яркости между соседними участками зрительного поля. Так, серый квадрат на черном фоне кажется светлее, чем такой же квадрат на светлом фоне (рис. 3).
Американские исследователи Х.К.Хартлайн и Ф.Ратлиф при помощи электрофизиологических методов обнаружили наличие тормозных взаимодействий между рецепторными элементами глаза пресноводного рачка limulus . Тормозное влияние, оказываемое рецептором А на рецептор Б, оказалось пропорциональным освещению А и пространственной близости обоих элементов. Это явление получило название латерального торможения.
В результате латерального торможения, оказываемого соседними ярко освещенными элементами, расположенный на краю слабоосвещенной области рецептор будет разряжаться с меньшей частотой, чем элементы, освещенные столь же слабо, но расположенные дальше от границы двух областей. Напротив, рецептор, лежащий на краю ярко-освещенной области, будет разряжаться с большей частотой, чем рецепторы, расположенные в глубине этой области. Таким образом, благодаря латеральному торможению, картина возбуждений элементов сетчатки подчеркивает границы между областями различной яркости.
Тормозные взаимодействия были обнаружены в зрительных системах высокоорганизованных животных, включая обезьян. В то же время существуют данные о значительно более сложном, центральном происхождении явления контраста.
Так, например, на контраст влияет сознательная установка наблюдателя. Если знак, изображенный на рис. 4, воспринимать как две ла-
| Величковский Б . М ., Зинченко В , П ., Лурия А.Р . Восприятие цвета 117 |

Рис. 4. Влияние установки наблюдателя на яркостный контраст (по: Коффка К., 1935)
тинские буквы V, то наблюдается выраженный яркостный контраст: левая буква кажется более светлой, чем правая. Если же воспринимать этот знак как одну букву W, то контраст исчезает,
Наряду с только что рассмотренным одновременным контрастом известен также последовательный яркостный контраст. Он выступает в виде последовательных образов — зрительных ощущений света, продолжающихся некоторое время после окончания действия раздражителя.
Различают отрицательные и положительные последовательные образы. Первые возникают, если при нормальном освещении в течение примерно 30 с рассматривать ярко освещенный объект, а затем быстро заменить его равномерным полем нейтрального цвета с более низкой яркостью. В этом случае испытуемый некоторое время видит перед собой темное пятно, по форме отдаленно напоминающее объект. Если же объект освещается в темноте вспышкой света, то возникает положительный последовательный образ. Как правило, он исчезает значительно быстрее, чем отрицательный. <...>
Второй характеристикой апертуарных цветов является цветовой тон. Монохроматические, т.е. вызванные светом с одной длиной волны, красный, зеленый, желтый и другие цвета одинаковой видимости яркости различаются как раз по своему цветовому тону. Это качество цветовых ощущений связано прежде всего с длиной волны раздражителя.
При переходе от коротковолновых к длинноволновым электромагнитным колебаниям цветовой тон меняется следующим образом: раздражители с короткими длинами волн воспринимаются фиолетовыми, затем следует узкий участок чистого синего цвета, который кончается сине-зелеными тонами, далее находится узкая полоска чистого зеленого цвета, за которым следуют желто-зеленые тона, потом появляется чистый желтый цвет и, наконец, в длинноволновой области — желто-красные цветовые тона.
Таким образом, монохроматические цветовые тона переходят друг в друга, образуя непрерывный ряд. Этот ряд можно превратить в замк-
118
Тема 17. Экспериментальные исследования восприятия

нутый цветовой круг, если добавить к нему пурпурные (фиолетово-красные) цветовые тона, не являющиеся монохроматическими (рис. 5).
Зрительная система способна различать очень тонкие оттенки цветового тона. Общее число различных оттенков монохроматических тонов достигает 150—200. Минимальные разностные пороги, равные 1 нм, найдены в сине-зеленой (485 нм) и зеленовато-желтой (575 нм) частях спектра.
Если длина волны однозначно определяет цветовой тон, то обратное утверждение неверно. Одному и тому же цветовому тону соответствует бесчисленное множество различных комбинаций монохроматических раздражителей. Законы смешения цветов были открыты И.Ньютоном не позже 1692 г. Однако полностью их справедливость была доказана только в прошлом веке. Известны три таких закона.
1. Для каждого цветового тона существует дополнительный цветовой тон, смешение с которым в определенной пропорции дает ощущение одного из оттенков серого (нейтрального) цвета. Следующие пары цветов являются дополнительными:
красный (660 нм) — сине-зеленый (497 нм)
оранжевый (610 нм) — зелено-синий (494 нм)
желтый (585 нм) — синий (485 нм)
желто-зеленый (570 нм) — фиолетовый (430 нм).
Легко видеть, что дополнительные цветовые тона расположены примерно на противоположных концах диаметров цветового круга.
Величковский Б . М ., ЗинченкоВ . П ., Лурия А . Р . Восприятие цвета 119

Рис. 6. Пропорции красного (650 нм), зеленого (530 нм) и синего (460 нм) цветов, необходимые для получения всех цветовых тонов спектра
2. При смешении двух цветов, лежащих на цветовом круге ближе, чем дополнительные, цветовой тон смеси расположен между смешиваемыми цветами на соединяющей их прямой,
3. Одинаково выглядящие цвета, независимо от своего спектрального состава, дают при смешении одинаковые по цветовому тону смеси.
Наиболее важное следствие из законов смешения цветов заключается в том, что с помощью любых трех цветов, не являющихся дополнительными, можно получить любой цветовой тон. Тройки цветов, отвечающие этому требованию, называются основными цветами. К ним относятся, например, красный, синий и зеленый цвет. На рис. 6 показано, в каких пропорциях нужно брать монохроматические красный, зеленый и синий тона, чтобы получить все остальные цветовые тона видимого спектра.
Степень отличия некоторого цветового тона от нейтрального тона, равного с ним по светлоте, определяет третью и последнюю характеристику цветовых тонов — их насыщенность. Физическим коррелятом насыщенности является «зашумленность» спектрального состава света электромагнитными колебаниями с другими длинами волн.
Насыщенность зависит также от яркости стимулов. Она максимальна для средних уровней освещенности и падает как при увеличении, так и при уменьшении яркости, вплоть до полного обесцвечивания раздражителей. Синие, красные и пурпурные цвета кажутся сильно насыщенными и остаются насыщенными даже при низких уровнях яркости, желтые и зелено-желтые становятся относительно насыщенными при больших яркостях.
Как и в случае видимой яркости, существуют цветовая адаптация и цветовой контраст. Они выражаются в общем или локальном уменьшении воспринимаемой величины насыщенности цветового тона при его
120
Тема 17. Экспериментальные исследования восприятия
белый длительном рассматривании и в одновре-
|
|

менном возрастании видимой насыщенности дополнительного цвета.
| с-з |
Цветовой последовательный контраст проявляется в виде цветовых последовательных образов. Например, длительная фиксация красного квадрата приводит к тому, что наблюдатель видит затем некоторое время перед собой зеленоватый квадрат, смещающийся вместе с движениями глаз. Еще И.В.Гёте обратил внимание на то, что цвета последовательных образов несколько отличаются от дополнительных, по сравнению с которыми они сдвинуты к краям спектра.
| ЧЕРНЫЙ Рис, 7. Цветовое тело (см. текст) |
Все многообразие апертурных цветов, определяемых тремя рассмотренными характеристиками, можно представить в виде простой пространственной модели — цветового тела (рис. 7). Оно представляет собой двойную коническую пирамиду, по вертикальной оси которой происходит увеличение светлоты цвета. Каждое горизонтальное сечение цветового тела является цветовым кругом для данного уровня видимой яркости. Насыщенность цветового тона уменьшается при движении по радиусу цветового круга, в центре которого находится нейтральный серый цвет. Уменьшение радиусов цветовых кругов на концах цветового тела объясняется уменьшением насыщенности цветов при низких и высоких уровнях видимой яркости.
Вследствие того, что все цветовые тона, включая нейтральные, могут быть получены с помощью смешения трех основных цветов, на практике для описания цветов пользуются цветовым телом, сечение которого представляет собой не круг, а треугольник. На вершинах этого цветового треугольника находятся три основных цвета: красный, зеленый, синий.
На рис. 8 показан цветовой треугольник, принятый Международной осветительной комиссией (МОК) в качестве эталонного. Условные коэффициенты на осях X и Y определяют координаты каждого цвета внутри цветового треугольника. Укажем координаты цветности некоторых цветов;
| X | Y | |
| красный | 0,67 | 0,33 |
| зеленый | 0,21 | 0,71 |
| синий | 0,14 | 0,08 |
| голубой | 0,16 | 0,31 |
и т.д.
| Величковский Б . М .. Зинченко В . П ., Лурия А . Р . Восприятие цвета 121 |

О 0,1 В т 0,20 0,30 О.Ш 0, М 0. S 0 0,70 D .80 X
Рис. 8. Цветовой треугольник МОК (см.текст)
Существуют две классические теории цветовых ощущений, называемые трехкомпонентной теорией и теорией противоцветов.
Первые идеи о трехкомпонентности цветового зрения были высказаны М.В.Ломоносовым в его «Слове о происхождении света, новую теорию о цветах представляющем, июля 1 дня 1756 года говорением». Эта теория была детально разработана в XIX в. английским физиком Т.Юнгом и Г.Гельмгольцем.
Теория основана на предположении, что число различных рецепторов цвета в сетчатке должно быть невелико. Действительно, если предположить, что для каждого из воспринимаемых нами оттенков существует специальный рецептор, то в условиях монохроматического освещения работало бы меньше одного процента рецепторов и зрение должно было бы резко ухудшаться. Простые наблюдения показывают, что этого не происходит.
122 Тема 17. Экспериментальные исследования восприятия
Так как все цвета могут быть получены с помощью смешения трех основных, то было сделано предположение, что в сетчатке существуют три типа рецепторов, чувствительных к синему (фиолетовому), зеленому и красному цветам.
Альтернативную теорию выдвинул Э.Геринг (1878)1. В основу теории противоцветов легли данные о подробно изученных им явлениях контраста, а также некоторые психологические наблюдения. Так, большинство людей выделяют в качестве основного «главного») цвета, помимо красного, зеленого и синего цветов, также и желтый. Э.Геринг считал, что в сетчатке находятся три цветочувствительных субстанции. Их разложение приводит к восприятию белого, зеленого и желтого цветов, а восстановление соответственно к восприятию черного, красного и синего цветов. Обе теории долгое время противопоставляли друг другу. Одной из областей, в которой их сторонники искали подтверждения своих взглядов, было исследование различных аномалий цветового зрения.
Нарушения цветового зрения встречаются приблизительно у 8% мужчин и 0,5% женщин. Эти нарушения, по крайней мере отчасти, являются наследуемыми. Было бы неправильно называть этих людей цветослепыми, так как лишь один исключительно редкий вид расстройства цветового зрения связан с полной неспособностью различать цветовые тональности. Люди с такими недостатками называются монохроматами. В этом случае все длины волн и все смеси различаются исключительно по своей светлоте.
Значительная часть нарушений цветового зрения связана с затруднениями в дифференциации красного и зеленого цветов. Особые трудности эти люди испытывают при различении таких цветов, как голубой и розовый. Трехкомпонентная теория, исходящая из существования трех первичных видов рецепторов, объясняет эту аномалию выпадением рецепторов, чувствительных к красному или зеленому цветам. И действительно, было обнаружено, что существуют две разновидности красно-зеленой слепоты. В опытах на получение желтого цвета одним из этих цветоаномалов требовалось гораздо больше красного, а другим — зеленого цвета, чем людям с нормальным зрением. Первая разновидность — нечувствительность к красному — была названа протанопией, а вторая — нечувствительность к зеленому — дейтеропией.
В пользу трехкомпонентной теории говорит, в свою очередь, существование слепоты на синий цвет, который путается в этом случае с зеленым. Это нарушение встречается почти столь же редко, как и полная цветослепота.
В то же время более детальные исследования показали, что красно-оранжево-желто-зеленая часть спектра преобразуется в восприятии цветоаномала не в оттенки зеленого (протанопия) или оттенки красного (дейтеропия), а в оттенки желтого цвета. Можно предположить, таким
1 Некоторые замечания, предвосхищающие теорию Э.Геринга, можно найти у Леонардо да Винчи и И.В.Гёте.
Величковский Б . М ., Зинченко В . П ., Лурия А . Р , Восприятие цвета 123
образом, что красно-зеленая слепота представляет собой дихроматическое желто-синее зрение. Этот факт в большей степени соответствует теории противоцветов Э.Геринга.
Аргументом в пользу теории противоцветов служат полученные на нормальных людях данные о порядке исчезновения цветового тона при перемещении стимулов в периферическое зрение. В этом случае первыми одновременно исчезают красные и зеленые цвета, от которых остается только желтый оттенок. Желтый и синий цветовые тона воспринимаются в более широкой области зрительного поля. Эти эффекты следует учитывать при использовании цветовой индикации.
Многочисленные доказательства как в пользу трехкомпонентной теории, так и в пользу теории противоцветов позволили Л.А.Орбели предположить, что справедливы обе теории. Однако каждая из них описывает закономерности переработки информации о цвете на различных уровнях зрительной системы. В последние годы детальное обоснование этой точки зрения было проведено американскими исследователями Л.М.Гурвичем и Д.Джексон. На рис. 9 показана разработанная ими схема отношений между тремя светочувствительными субстанциями и четырьмя реципроктными процессами, лежащими в основе цветового зрения. Недавно были получены прямые нейрофизиологические доказательства справедливости этой модифицированной теории.
Прежде всего удалось показать, что в сетчатке действительно имеются три светочувствительных вещества. Один из самых тонких опытов в этой области был проведен английскими исследователями П.К.Брауном и Дж.Уолдом.

Рис. 9. Упрощенная схема взаимоотношений между светочувствительными
веществами и тремя парами противоположных процессов: сине-желтым,
красно-зеленым и бело-черным (по: Гурвич JIM., Джемсон Д., 1966)
124
Тема 17. Экспериментальные исследования восприятия

Рис. 10. Регистрация импульсов одиночных
ганглиозных клеток сетчатки (по: Гранит Р., 1955)
После избирательной адаптации к соответствующим дополнительным
цветам выявлено наличие рецепторов для синего (I), зеленого (II) и
красного (III) цветов
В их экспериментах миниатюрный пучок монохроматического света проецировался через зрачок на одиночные колбочки сетчатки испытуемого и с помощью микроспектрофотометра измерялось количество отраженного и вернувшегося через зрачок света. Было установлено, что существуют три типа колбочек, имеющих максимумы поглощения при 450, 525 и 535 нм.
Электрофизиологические опыты с микроэлектродной регистрацией активности ганглиозных клеток сетчатки также говорят о существовании трех типов цветовых рецепторов. Шведский физиолог Р.Гранит показал, что возрастание активности нейронов возникает в ответ на освещение сетчатки синим, зеленым или красным светом (рис. 10).
Если исследования механизмов цветового зрения на уровне сетчатки подтверждают трехкомпонентную теорию, то исследования на более высоком уровне латерального коленчатого тела говорят в пользу теории противоцветов. Целый ряд работ, среди которых можно отметить исследования американского физиолога Р.Л.де Валуа и Е.Н.Соколова, показали, что на этом уровне наблюдаются реакции оппонентного типа. Например, были найдены нейроны, увеличивающие активность в ответ на освещение сетчатки красным светом и уменьшающие ее в ответ на зеленый свет. Наряду с такими «красно-зелеными» элементами были найдены такке «желто-синие» и «бело-черные» нейроны.
Таким образом, классические теории цветового зрения не исключают, а дополняют друг друга.
Г . Глейтман ,
А . Фридлунд
Д , Райсберг
ИССЛЕДОВАНИЕ ОЩУЩЕНИЙ1
Развитие психологических методов, а также разнообразных психологических техник вооружило психологию серьезным инструментарием для изучения ощущений. Мы в основном остановимся на зрении, поскольку есть основания полагать, что именно эта модальность доминирует в перцептивной системе человека. Но сначала сделаем краткий обзор других видов ощущений, снабжающих нас информацией об окружающем мире и о нашем положении в нем.
Кинестетика и вестибуляторная система
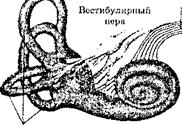
Эти чувства информируют организм о его собственных движениях и местоположении в пространстве. Перемещения костей скелета (т.е. движения рук, ног, шеи и т.д.) отслеживаются с помощью кинестетики (собирательное название всей информации, поступающей от рецепторов, расположенных в мышцах, связках и суставах). Другая группа рецепторов сигнализирует обо всех движениях головы — произвольных или возникающих в результате воздействия внешних сил. Эти рецепторы локализованы в трех полукружных каналах, расположенных в преддверии внутреннего уха (рис. 1). Внутри этих каналов находится вязкая жидкость, которая приходит в движение при изменении положения головы. От этого перемещения деформируются волосковые клетки, которые расположены на концах каждого канала. Изменяя свою форму, эти волосковые клетки возбуждают нервный импульс. Совокупность импульсов от
1 Глейтман Г., Фридлунд А., Райсберг Д, Основы психологии. СПб.: Речь, 2001. С. 211-212, 216-226.
126
Тема 17, Экспериментальные исследования восприятия
|
|

___ Слуховой
* '* \ нерв
Полукружные каналы
Б
А
Рис. 1. Вестибулярная система: А — местоположение внутреннего уха. Этот парный орган расположен с обеих сторон черепа. Остальные структуры внутреннего уха обслуживают чувство слуха1; Б — изображение вестибулярного аппарата2
каждого из каналов дает информацию о причинах и амплитуде движения головы.
Одной из жизненно важных функций системы полукружных каналов является обеспечение устойчивой «платформы» для зрения. В процессе ходьбы мы все время совершаем движения головой. Для того чтобы компенсировать эти бесконечные движения, нашим глазам приходится совершать равнозначные перемещения. Такое приспособление происходит благодаря наличию вестибулярной системы, тесно связанной с мозжечком в заднем отделе головного мозга, который нивелирует каждый поворот головы равным и пртивонаправленным движением глаз. Эти перемещения запускаются благодаря сообщениям, приходящим из трех полукружных каналов, которые затем передаются соответствующим мышцам каждого глаза. Таким образом, зрительная система является совершенно стабильной. <...>
Кожные чувства
Стимуляция кожных рецепторов информирует организм о том, что находится непосредственно вблизи тела. Неудивительно, что чувствительность кожи особенно сильна в тех частях тела, с помощью которых
1 См.: Krech D., Crutchflld R. Elements of psychology. N. Y.: Knopf, 1958.
2 См.: Kalat J.W. Biological psychology. Belmont, Calif.: Wadsworth, 1984.
Глейтман Г . Фридлунд А, Райсбврг Д . Исследование ощущений 127
люди изучают окружающий мир «без посредников»: это ладони и пальцы, губы и язык. Получаемые с помощью этих «разведчиков» ощущения сказываются на организации проекционной зоны коры головного мозга, отвечающей за телесный опыт. Как мы уже знаем, существующее распределение кортикального пространства крайне неравномерно, причем львиную долю поверхности коры занимают участки, отвечающие за такие чувствительные части нашего организма, как лицо, рот и пальцы
Сколько существует кожных чувств? Аристотель полагал, что все ощущения, связанные с кожей, можно свести к одному — прикосновению. Но в наши дни исследователи выделяют по меньшей мере четыре различных кожных чувства: давление, тепло, холод и боль. Некоторые удлиняют этот список и включают в него другие ощущения. Среди них — ощущения вибрации, щекотки и почесывания. Каким же образом такие разные виды сенсорной информации кодируются в нашей нервной системе?
Ответ на этот вопрос впервые дал психолог Дж.Мюллер (1801 — 1858). Он провозгласил доктрину специфических нервных энергий — заявил о том, что различия в качествах ощущений вызваны различиями нервных структур, возбуждаемых разными стимулами. Например, какая-то одна нервная структура может дать сигнал «тепло», а другая — «холодно», и т.д.
Верно ли это? Без сомнений можно ответить: «Да». Есть все основания полагать, что разнообразные ощущения давления, к примеру, вызваны специфическими рецепторами, находящимися в коже <...>. Некоторые из этих рецепторов расположены вокруг волосяных фолликул в коже; они реагируют на движения волоса. Другие существуют в виде капсул, которые легко сжимаются при малейшей деформации кожи. Капсулы одного вида реагируют на постоянную вибрацию, капсулы другого вида — на внезапные раздражения кожи, третий вид капсул отзывается на равномерное надавливание. Очевидно, что существует не один вид тактильных рецепторов, а несколько.
Гораздо меньше нам известно о рецепторной системе, воспринимающей тепло, холод и боль. Возможно, за некоторые из этих ощущений отвечают свободные нервные окончания, не имеющие специфического органа-эффектора и находящиеся в коже. Раньше принято было считать, что эти свободные нервные окончания передают информацию о тепле, холоде и боли, но, возможно, некоторые из них являются дополнительными рецепторами давления1.
1 См.: Sherrick СЕ ,, Cholewiak R.W. Cutaneous sensitivity // K.R.Boff, L.Kaufman, J.P.Thomas (Eds.). Handbook of perception and human performance. N. Y.: Wiley, 1986. Ch. 12.
128
Тема 17. Экспериментальные исследования восприятия
Чувство вкуса
Чувство вкуса играет роль стража пищеварительной системы организма, поставляя информацию о тех веществах, которые стоит или не стоит пускать внутрь. Его задача — отвадить яды и пригласить пищу. ¥ большинства обитающих на суше млекопитающих эту функцию выполняют специальные рецепторные органы, заполненные вкусовыми сосочками, которые очень чувствительны к растворенным в воде химическим веществам. В среднем у человека около 10000 таких вкусовых сосочков. Большая часть их находится на кончике языка, но некоторые расположены и на других участках полости рта. Нервные волокна от этих рецепторов передают сообщение к головному мозгу: сначала — в продолговатый мозг, а далее — в таламус и кору.
Вкусовые ощущения
Многие исследователи полагают, что существует четыре основных вкуса: кислый, сладкий, соленый п горький. На их взгляд, все остальные вкусовые ощущения образовались от смешения этих первичных вкусов. Так, грейпфрут на вкус кисло-горький, а лимонад — кисло-сладкий.
По-видимому, каждый из этих основных вкусов играет свою неповторимую биологическую роль. Большинство организмов привлекает сладкий вкус. Вероятно, это происходит вследствие того, что многие питательные вещества содержат в себе в той или иной форме сахар. А поскольку большинство ядовитых веществ имеет горький вкус, на ранних стадиях развития человечество выработало защитные рефлексы против их поглощения — отрыжку, рвоту, — которые координируются задними отделами головного мозга1.
Какие же стимулы вызывают эти четыре основных вкуса? До сих пор у нас нет исчерпывающего ответа на этот вопрос. Нам точно известно, что вкус кислого возникает благодаря работе рецепторов, чувствительных к кислоте, а соленый вкус — это результат чувствительности некоторых рецепторов к наличию натрия. Со сладким и горьким вкусами дело обстоит сложнее. И тот и другой, как правило, вызываются сложными органическими молекулами, но до сих пор не существует четко сформулированных правил, описывающих взаимосвязь между молекулярной структурой и получаемым вкусом. Сладкий вкус вызывают производные различных Сахаров, но также и искусственного подсластителя — сахарина, который по своему химическому составу сильно от-
1 См.: Shepherd G.M. Discrimination of molecular signals by the olfactory receptor neuron // Neuron. 1994. 13. P, 771-790; Shepherd G.M. Neurobiology. N. Y.: Oxford University
Press, 1994.
Глейтман Г. , Фридлунд А, Райсберг Д . Исследование ощущений 129
личается от Сахаров. Горький вкус вызывается разнообразными химическими веществами, что порождает гипотезу о существовании нескольких видов рецепторов горького вкуса.
Вкус и сенсорное взаимодействие
Вкусовые ощущения могут послужить иллюстрацией к закономерности, которой подчинены все модальности. Эту закономерность мы назовем сенсорным взаимодействием. Она описывает тот факт, что реакция сенсорной системы на любой стимул обычно зависит не только лишь от одного этого стимула. На реакцию влияют и другие стимулы, воздействующие на систему в данный момент или взаимодействовавшие с ней некоторое время назад.
Один из вариантов сенсорного взаимодействия можно наблюдать довольно часто. Предположим, что вкусовой стимул предъявляется непрерывно в течение 15 с или дольше. В результате произойдет адаптация — феномен, обнаруженный во всех сенсорных системах. Например, если язык постоянно стимулировать чем-то соленым, чувствительность ко всему соленому понизится. Аналогично, после долгой и непрерывной дегустации раствора хинина он будет казаться все менее и менее горьким. Однако этот адаптационный процесс обратим. Если прополоскать рот и не воспринимать никаких вкусовых раздражителей, скажем, в течение минуты, первоначальная вкусовая чувствительность будет полностью восстановлена.
В другом виде взаимодействия адаптация к одному вкусу может привести к усилению другого, этот эффект иногда рассматривают как форму контраста. К примеру, адаптация к вкусу сахара заставляет кислоту казаться кислее, чем это было раньше1. Точно так же, адаптировавшись к соленому раствору, мы почувствуем, что обыкновенная водопроводная вода стала как будто более кислой или горькой; а адаптировавшись к сладкому, скажем, что эта же вода определенно стала более горькой2,
Обоняние
Мы обсудили сенсорные системы, которые дают информацию о предметах и событиях вблизи нас: о движении и местоположении наших тел, об ощущениях нашей кожи, о том, что мы только что с аппетитом
1 См.: Kuznicki J.T, McCutcheon N.B, Cross enhancement of the sour taste of single
human taste papillae .// Journal of Experimental Psychology. 1979, 198. P. 68-89.
2 См.; McBurney DM., Shick Т . Н . Taste and water taste of twenty-six compounds for
man // Perception and Psychophysics. 1971, 10. P. 249-252.
130
Тема 17. Экспериментальные исследования восприятия
съели. Но ведь мы получаем информацию и о том, что находится далеко от нас. Человек обладает тремя главными сенсорными системами, реагирующими на дальние стимулы: это — обоняние, слух и зрение.
Обоняние как дистантное чувство
Как сенсорная система, реагирующая на дальние стимулы, обоняние жизненно важно для многих видов животных. Обоняние — первейшее средство для поиска пищи, обнаружения хищников и сородичей. Запах играет меньшую роль в человеческом сообществе, в стадах приматов и в птичьих стаях. Предки всех этих созданий покинули кишащую запахами поверхность земли, дабы подняться к деревьям, а в этой древесной среде более значимыми стали другие чувства, особенно зрение. Мы можем проверить это контрастное для многих видов положение дел, используя психофизические методы. Окажется, например, что чувствительность собак к запахам приблизительно в тысячу раз превосходит чувствительность людей1.
По сравнению с большинством обитателей поверхности земли, человеческие создания весьма убоги в области обоняния. Но это вовсе не означает, что в человеческой жизни нет места запахам. Они предупреждают нас о надвигающейся опасности (когда, например, мы чувствуем запах газа), они добавляют много приятных моментов тогда, когда мы едим что-нибудь вкусное; запахи — основа всей парфюмерно-дезодорантной индустрии. Согласно некоторым опубликованным данным, они даже помогают продавать чемоданы и подержанные автомобили: пластмассовые портфели, пропитанные искусственным запахом кожи и далеко не новые машины, «спарфюмированные» под только что сошедшие с конвейера, имеют большую рыночную стоимость2. Кроме того, запах играет роль и при узнавании людей. В одном из исследований ученый-психолог попросил нескольких мужчин и женщин в течение одних суток носить футболки, не принимая душ и не пользуясь парфюмерией и дезодорантами. По истечении 24 часов каждая (нестиранная) футболка была упакована в отдельный пакет. Затем всех участников исследования попросили понюхать то, что находилось в трех пакетах, не заглядывая внутрь. В одном из них была его (или ее) футболка, во втором — футболка другого мужчины, в третьем — та, которую носила какая-то другая женщина. Около 75% испытуемых смогли найти свою футболку, опираясь
1 См.: Marshall DA„ Moult on D.G. Olfactory sensitivity to a-ionine in humans and dogs
// Chemical Senses. 1981. 6. P. 53-61; Cain W.S. Olfaction // R.C. Atkinson, R.J. Herrnstein,
G. Lindzey, R.D. Luce (Eds.). Stevens' handbook of experimental psychology. N. Y.: Wiley,
1988. У. 1. Perception and motivation, rev. ed. P. 409-459.
2 См.: Winter R. The smell book: Scents, sex, and society. Philadelphia: Lippincott,
1976.
Глейтман Г ., Фридлунд А, Райсберг Д . Исследование ощущений 131
лишь на запах, и, кроме того, им удалось правильно определить, мужчина или женщина носили каждую из двух других футболок1.
Обоняние как контактное чувство
Обоняние — такое чувство, которое дает нам очень важную информацию не только о предметах, находящихся на большом расстоянии, но и близко от нас. Возьмем, к примеру, то, что находится у нас во рту2. Мы можем почувствовать запах котлет, лежащих на нашей тарелке, но также мы можем распробовать их вкус, положив котлету в рот. Этот вкус — как, впрочем, и все вкусы вообще — зависит, в основном, от нашего обоняния. Потому что то, что мы привыкли называть «вкусом» еды, редко является результатом работы лишь вкусовых рецепторов; почти всегда это комбинация вкуса, текстуры, температуры и — что важнее всего — запаха. Когда у нас сильный насморк, нам кажется, что еда совершенно лишена вкуса. И хотя в такие моменты мы все еще различаем четыре основных вкуса — аромат потерян и еда кажется безвкусной. Если исчез запах, мы уже не сможем отличить уксус от тонкого красного вина или яблоко от луковицы. Для гурмана, шеф-повара или дегустатора вин чувствительный нос иногда гораздо более важен, чем чувствительный язык.
Слух
Слух представляет собой реакцию на такую физическую величину, как давление, и с этой точки зрения является близким родственником кожных чувств. Однако, в отличие от тактильных ощущений, слуховые ощущения информируют нас о том, что давление изменяется благодаря событиям, происходящим на расстоянии многих метров от нас.
Звук
Что представляют собой слуховые стимулы? В окружающем нас мире все время происходят какие-то физические движения — зверь прошмыгнул в зарослях кустарника или колеблются голосовые связки у вашего соседа. Такое движение активизирует частицы воздуха, находя-
1 См.: Russell M . J . Human olfactory communication // Nature. 1976. 260. P. 520-22;
McBurney D.H., Levlne J.M., Cavanaugh Р . Я . Psychophysical and social ratings of human
body odor // Personality and Social Psychology Bulletin. 1977. 3. P. 135-138.
2 См.: Rosin P. Human food selection: The interaction of biology, culture, and individual
experience // L.M. Barker (Ed.). The psychology of human food selection. Westport, Conn.:
AVI Publ. Co., 1982.
132
Тема 17. Экспериментальные исследования восприятия
щиеся вокруг двигающегося объекта, а они, в свою очередь, «толкают» другие частицы, которые сообщают этот импульс дальше. Само перемещение частиц очень незначительно (около одной миллионной доли сантиметра) и непродолжительно (частица возвращается в свое исходное положение спустя несколько тысячных секунды), но этого действия достаточно для того, чтобы создать кратковременный импульс, расходящийся во все стороны от движущегося объекта наподобие кругов на воде, в которую бросили камень.
Даже если движение длится очень недолго, оно порождает серию колебаний в воздухе. Когда звуковые волны достигают уха, они инициируют дальнейшие микроизменения и в конце концов срабатывают слуховые рецепторы. Затем рецепторы запускают различные нейрональные реакции, которые, в свою очередь, достигают головного мозга и заставляют нас переживать слуховые ощущения.
Звуковые волны характеризуются амплитудой и частотой. Амплитуда описывает давление, сообщаемое каждой частицей воздуха своей соседке. Это давление колеблется от минимального до максимального по мере движения звука. Обычно та амплитуда, которую мы стараемся измерить, соответствует максимальному уровню давления, возникающему на гребне звуковой волны. Частота волны описывает частоту возникновения ее гребней. Сколько времени проходит между одним из гребней волны и следующим за ним? Этот интервал называют длиной звуковой волны. Хотя более обобщенно звуковые волны описываются частотой, которая определяется количеством ее гребней (пиков) в секунду. Поскольку скорость звука в какой-либо среде постоянна, частота обратно пропорциональна длине волны (рис. 2).
Длина волны

Рис. 2. Стимул для слуха
Колеблющийся предмет определенным образом толкает окружающие его молекулы; затем эта пульсация распространяется, как круги по воде, в которую бросили камень. Чтобы описать это пример, понадобится измерить давление воздуха в единичной точке пространства. Давление звука колеблется, как показано на этом рисунке. Максимальное давление определяется амплитудой звуковой волны; интервал между пиками давления определяет длину волны
Глейтман Г., Фридлунд А. , Райсберг Д . Исследование ощущений 133
И амплитуда, и частота суть физические характеристики самой звуковой волны, но они достаточно легко сопоставляются с такими психологическими величинами, как громкость и высота звука. Проще говоря, звук нам будет казаться громче по мере увеличения его амплитуды и выше — по мере возрастания частоты.
Амплитуда и громкость звука. Диапазон звуковой амплитуды, воспринимаемой человеком, настолько огромен, что ученые решили измерять эту величину с помощью логарифмической шкалы; поэтому гром-
Таблица 1
Громкость различных звуков
| Звук | Громкость звука, дБ |
| Запускаемый космический корабль (расстояние 50 м) | 180 |
| Самая громкая рок-группа | 160 |
| Болевой порог (приблизительный) | 140 |
| Близкий гром; средняя рок-группа | 120 |
| Крик | 100 |
| Шумный автомобиль | 80 |
| Нормальный разговор | 60 |
| Шепот | 20 |
| Шорох листьев | 10 |
| Слуховой порог | 0 |
Таблица 2 Частоты некоторых музыкальных звуков
| Звук | Частота звука, Гц |
| Самая высокая нота рояля | 4214 |
| Самая высокая нота флейты-пикколо | 3951 |
| Самый высокий звук сопрано | 1152 |
| Самый высокий звук альта | 640 |
| Среднее «до» | 256 |
| Самый низкий звук баритона | 96 |
| Самый низкий звук баса | 80 |
| Самая низкая нота контрабаса | 29 |
| Самый низкий звук рояля | 27 |
| Самый низкий звук органа | 16 |
134
Тема 17. Экспериментальные исследования восприятия
кость звука измеряется в децибелах (табл. 1). В психологическом смысле воспринимаемая громкость возрастает в два раза, если громкость звука увеличивается на 10 дБ1.
Частота и высота звука. Частота звука обычно измеряется количеством полных волновых циклов в секунду, или герцами (по имени германского физика Генриха Герца). Частоты, соответствующие различным музыкальным звукам, показаны в табл. 2. Молодые взрослые люди могут слышать звуки частотой от 20 до 20000 Гц, причем максимальная чувствительность наблюдается для звуков из средней части этого диапазона. По мере старения человека его звуковая чувствительность ухудшается, особенно это относится к высоким частотам.
Возникновение ближних стимулов
У млекопитающих слуховые рецепторы расположены глубоко внутри уха в органе, по форме своей напоминающем улитку (он так и называется — улитка). Чтобы достичь улитки, звуку приходится пробираться нелегким путем. Наружное ухо собирает звуковые волны из воздушной среды и направляет их к барабанной перепонке — упругой мембране, находящейся в конце слухового прохода. Звуковые волны заставляют барабанную перепонку колебаться, а эти колебания, в свою очередь, передаются на пластинку овального отверстия, разделяющую среднее ухо и внутреннее ухо. Операция передачи осуществляется тремя крошечными косточками, имеющими общее название «слуховые косточки». Колебания барабанной перепонки воздействуют на первую косточку, которая, начав двигаться, передает это движение второй косточке, а она — третьей, которая и завершает эту цепочку, сообщая «рисунок» колебания прикрепленной к ней пластинке овального отверстия. Колебание пластинки овального отверстия порождает колебания жидкости, находящейся внутри улитки, вызывая реакцию рецепторов
Зачем существует такой окольный путь передачи звука? Звуковые волны приходят к нам по воздуху, и ближний стимул, характерный для слуха, представляет собой кратковременный перепад воздушного давления. Но внутреннее ухо заполнено улиточной жидкостью (перилимфой). Поэтому, чтобы мы что-нибудь услышали, изменения воздушного давления должны вызвать изменения давления жидкости. Известно, что жидкость привести в движение гораздо труднее, чем воздух. Чтобы решить данную проблему, передающие давление волны должны быть каким-то образом усилены на пути к рецепторам; работу, связанную с их усилением, и выполняют различные части слухового органа. Например,
1 См.: Stevens S.S. The measurement of loudness // Journal of the Acoustical Society of America. 1955. 27. P. 15-19.
Глейтман Г ., Фридлунд А., Райсберг Д . Исследование ощущений 135
слуховые косточки выполняют функцию рычагов, используя рычажную силу для увеличения звукового давления. А барабанная перепонка примерно в двадцать раз больше по площади, чем пластинка овального отверстия, на которую воздействуют слуховые косточки. В результате довольно незначительная сила звуковых волн, воздействующих на барабанную перепонку, превращается в гораздо более внушительную силу, оказывая давление на меньшую по площади пластинку овального отверстия.
Преобразования в улитке
Почти по всей своей длине улитка разделена на верхнюю и нижнюю части при помощи нескольких структур, включая основную мембрану. Сами слуховые рецепторы называются волосковыми клетками. Эти клетки — в каждом ухе их примерно по 15 тысяч — расположены между основной мембраной и другими мембранами, находящимися выше <...>.
Перемещение пластинки овального отверстия вызывает изменение давления в улиточной жидкости, что, в свою очередь, заставляет колебаться основную мембрану. Колеблющаяся основная мембрана деформирует волосяные клетки, и, таким образом, самый непосредственный стимул воздействует на рецепторы. Каким образом движения волосяных клеток вызывают слуховое ощущение? Основной момент в данном вопросе — это восприятие высоты звука, а сенсорные качества высоты зависят от частоты звуковой волны.
Участки основной мембраны и высота звука. Согласно локализаци-онной теории высоты, впервые предложенной Германом Гельмгольцем (1821 —1894), различные участки основной мембраны реагируют на звуки разной частоты. А нервная система, в свою очередь, способна определить высоту звука по тому, в какой части основной мембраны колебание было более сильным. Стимуляция волосковых клеток одного конца основной мембраны приводит к ощущению высокого звука, а стимуляция волосковых клеток, находящихся на другом ее конце, вызывает ощущение низкого звука.
Постулат Гельмгольца был проверен в серии классических экспериментов Георга фон Бекеши (1899—1972), чьи работы, посвященные изучению слуха, принесли автору Нобелевскую премию в 1961 г. Работая с препаратами улиток людей и животных, Бекеши отделил часть стенки улитки таким образом, что смог наблюдать под микроскопом функционирование основной мембраны в моменты колебания пластинки овального отверстия, которое он раздражал с помощью электрического тока. Он обнаружил, что такая стимуляция приводит к волнообразной вибрации основной мембраны (рис. 3). Когда он стал варьировать частоту колебаний стимула, пики деформации начали возникать в разных областях мембраны, как и предполагал Гельмгольц. При высоких час-
136
Тема 17. Экспериментальные исследования восприятия
тотах пики обнаруживались ближе к овальному отверстию, а по мере снижения частоты этот пик смещался все ближе и ближе к верхушке улитки1. Получается, что высокие и низкие частоты воздействуют на разные волосковые клетки, в результате чего стимулируются различные слуховые нервные волокна и, значит, — различные участки головного мозга.
Частота звука и частота возбуждения нервных волокон. По мере того как снижается частота стимула, деформируемый участок основной мембраны становится все шире и шире. При частоте ниже 50 Гц волна, вызванная стимулом, деформирует почти равномерно всю поверхность мембраны2. Однако мы способны различать и звуки частотой 20 Гц, так что локализационная теория не описывает картину полностью. По-видимому, нервная система обладает еще каким-то арсеналом средств, помимо поверхности основной мембраны, для различения высоты звука.
Высоту звука можно определять с помощью теории частоты, которая основана на разнообразии частот передачи нервных импульсов по

Рис. 3. Деформация основной мембраны под действием звука: А — мембрана схематически изображена как простой прямоугольный лист бумаги. На самом деле, конечно же, она гораздо тоньше и свернута в спираль; Б — взаимосвязь между частотой звука и расположением пиков деформации основной мембраны. Пик деформации может располагаться на различных расстояниях от стремечка (третьей косточки, которая приводит в движение мембрану, ударяя по пластинке овального отверстия). Как показывает рисунок, чем выше частота звука, тем ближе к стремечку будет находиться этот пик3
1 См.: Bekeshi G. von. The ear // Scientific American. 1957. 197. P. 66-78.
2 См.: KhannaS.M., Leonard D.G.B.Basilar membrane tuning in the cat cochlea //
Science, 1982. 215. P. 305-306; Hudspeth A.J. How the ear's works work // Nature. 1989.
341. P. 397-404.
8 См.: Lindsey P.H., Norman DA. Human information processing. N. Y.: Academic Press, 1977; Coren S., Ward L.M. Sensation and perception. San Diego, Calif.: HarcourtBrace Jovanovich, 1989.
Глейтман Г ., Фридлунд А ., Райсберг Д . Исследование ощущений 137
слуховому нерву. Для частот ниже 50 Гц частота звукового стимула может быть преобразована непосредственно в соответствующее количество нервных импульсов в секунду. Затем эта информация поступает в вышележащие нервные центры, которые интерпретируют ее как определенную высоту звука.
Современные исследования показывают, что в восприятии высоты звука участвуют оба этих механизма. По-видимому, высокие частоты кодируются в зависимости от местоположения вызванного ими пика на поверхности основной мембраны, а низкие — в зависимости от частоты нервных импульсов. Местоположение пика играет незначительную роль в восприятии частот ниже 500 Гц, а частота импульса почти не влияет на восприятие звуков, частота которых превышает 5000 Гц. В среднем диапазоне, между 1000 и 5000 Гц, действенны оба механизма, и здесь высоты различаются очень точно1,
Дата добавления: 2020-01-07; просмотров: 170; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!