Микрофибриллы, или промежуточные микрофиламенты.
Это белковые струткуры. Толщиной 10 нм. Не ветвятся, часто располагаются слоями. Их белковый состав различен в разных тканях. В эпителии в состав микрофибрилл входит кератин. В мышечных клетках (кроме миоцитов сосудов) – белок десмин. Различных клетках мезенхимногшо происхождения ( фибробласты и др) – белок виментин.
Функция:
1. Опорно – каркасная, но они не так лабильны как микротрубочки.
Клеточный центр.
Это видимая в световой микроскоп структура (на границе его разрешающей способности), но ее тонкое строение можно изучить только с помощью электронного микроскопа.
Клеточный центр (центросома) состоит из центриолей и связанных с ними микротрубочек – центросферы. Термин «центриоли» был предложен Т.Бовери в 1895 году. Выявляются при окраске железным гематоксилином.
В интерфазной клетке присутствуют две центриоли, расположенные перпендикулярно друг к другу, образующие диплосому.
Центриоль представляет собой полый цилиндр. Стенка состоит из 9 триплетов микротрубочек, расположенных по окружности и соединенных ручками. Формула центриоли: (9х3)+0. Микротрубочки в центральной части центриоли отсутствуют. Вокруг каждой центриоли расположен бесструктурный, или тонковолокнистый матрикс. Каждый триплет микротрубочек связан со структурами сферической формы – сателлитами. От сателлитов расходятся в стороны микротрубочки, образуя центросферу.
|
|
|
При подготовке клетки к митотическому делению происходит удвоение центриолей. Он заключается в том, что две центриоли расходятся и около каждой вновь образуется дочерняя центриоль.
Функции:
1. Являются центром организации микротрубочек веретена деления.
2. Индуцирует полимеризацию тубулинов новой процентриоли, возникающей при ее дупликации.
3. Образование ресничек и жгутиков.
Органеллы специального значения.
Реснички и жгутики.
В световом микроскопе выглядят как тонкие выросты. Они имеются в некоторых клетках – сперматозоидах, эпителиоцитах трахеи и бронхов, семявыносящих путей мужчины, яйцеводах женщины.
В электронный микроскоп – цилиндрический вырост цитоплазмы диаметром 300 нм, покрытый плазматической мембраной. Внутри выроста расположена аксонема. Стенка аксонемы состоит из 9 пар микротрубочек, связанных «ручками». В центре аксонемы располагается пара центральных микротрубочек. Формула: (9х2)+2. В основании ресничек и жгутика в цитоплазме лежат мелкие гранулы – базальные тельца, сходные по своей структуре с центриолями. ((9х3)+0). Базальное тельце и аксонема структурно связаны между собой и составляют единое целое: две микротрубочки триплетов базального тельца являются микротрубочками дуплетов аксонемы. Основу микротрубочек составляет несократимый белок тубулин. Белок «ручек»- динеин – обладает АТФ-азной активностью: расщепляет АТФ,за счет энергии которой происходит смещение дуплетов микротрубочек друг по отношению к другу. Так совершаются волнообразные движения ресничек и жгутиков.
|
|
|
Функция:
- Специальные органоиды движения.
Миофибриллы находятся в мышечных клетках и миосимпластах. Являются органоидами сокращения.
Нейрофибриллы находятся в нейронах состоят из нейротубул и нейрофиламентов. Их функция - опорная и транспортная.
Тонофибриллы содержатся в эпителиоцитах. Участвуют в кератинизации.
Микроворсинки – увеличивают площадь всасывания. Содержатся, например, в эпителиоцитах кишечника, где участвуют в процессах пристеночного пищеварения.
Базальная складчатость – складки плазмолеммы, между которыми располагаются митохондрии, ориентированные перпендикулярно к базальной мембране.
Включения.
Включения цитоплазмы – необязательные компоненты клетки, возникающие и исчезающие в зависимости от метаболического состояния клетки.
|
|
|
Классификация включений:
1. Трофические включения – депонированные питательные вещества. К таким включениям относятся, например, включения гликогена, жира.
2. Секреторные включения- округлые образования различных размеров, содержащие биологически активные вещества, образующиеся в секреторных клетках
3. Экскреторные включения- подлежат выведению из клетки, поскольку состоят из конечных продуктов обмена.
4. Пигментные включения могут быть экзогенными (каротин, пылевые частицы, красители и др.) и эндогенными (гемоглобин, гемосидерин, билирубин, меланин, липофусцин). Наличие их в цитоплазме может изменять цвет ткани, органа временно или постоянно. Нередко пигментация ткани может служить диагностическим признаком.
Специальные включения- фагоцитированные частицы, поступающие в клетку путем эндоцитоза. Основными функциями ядра являются хранение и передача генетической информации, а также регуляция белкового синтеза. Составными компонентами ядра являются: ядерная оболочка, ядерный сок, хроматин, ядрышко и ядерный белковый матрикс.
Ядерная оболочка (нуклеолемма, кариолемма) построена по типу элементарной биологической мембраны. В электронный микроскоп она состоит из двух мембран – наружной и внутренней, между которыми располагается перинуклеарное пространство. В местах соединения этих мембран образуются отверстия – поры, обеспечивающие связь содержимого ядра с содержимым цитоплазмы. Количество пор непостоянно и зависит от активности клетки. Поры заполнены сложноорганизованными глобулярными и фибриллярными структурами, в совокупности формирующими так называемый комплекс поры. Он состоит из трех рядов периферических гранул, по 8 гранул в каждом ряду. Один ряд гранул располагается на уровне внутренней мембраны, второй – на уровне наружной, третий – между ними. В центре поры лежит непарная девятая гранула, от которой к периферическим гранулам идут тонкие белковые фибриллы, в совокупности образующие диафрагму. Если фибриллярные нити располагаются друг напротив друга – пора открыта, что и обеспечивает взаимодействие ядра и цитоплазмы. Основной функцией ядерной оболочки является барьерная – отграничение содержимого ядра от содержимого цитоплазмы.
|
|
|
Внешняя мембрана ядерной оболочки, непосредственно контактирующая с цитоплазмой клетки, имеет ряд структурных особенностей, позволяющих отнести ее к собственно мембранной системе эндоплазматической сети: на ней со стороны гиалоплазмы расположены многочисленные рибосомы, а сама внешняя ядерная мембрана может прямо переходить в мембраны эндоплазматической сети. Внутренняя мембрана связана с хромосомным материалом ядра.
Хроматин. Представляет собой зоны плотного вещества, хорошо воспринимающие различные красители, особенно основные. В интерфазе хроматин – это хромосомы, которые теряют в этот период свою компактную форму и разрыхляются (деконденсируются). В состав хроматина входит ДНК в комплексе с белками. В виде элементарных хромосомных фибрилл. В химическом отношении фибриллы хроматина представляют собой сложные комплексы дезоксирибонуклеопротеидов (ДНП), в состав которых входят ДНК и специальные хромосомные белки — гистоновые и негистоновые. Гистоны расположены по длине молекулы ДНК не равномерно, а в виде блоков. В один такой блок входят 8 молекул гистонов, образуя так называемую нуклеосом, поэтому хромосомная фибрилла имеет вид нитки бус или четок, где каждая бусина — нуклеосома Такие фибриллы толщиной 10 нм дополнительно продольно конденсируются и образуют основную элементарную фибриллу хроматина толщиной 25 нм.
В ядрах, кроме хроматиновых участков и матрикса, встречаются перихроматиновые фибриллы, перихроматиновые и интерхроматиновые гранулы. Они содержат РНК и встречаются практически во всех активных ядрах, представляют собой информационные РНК, связанные с белками, — рибонуклеопротеиды (информосомы). Матрицами для синтеза этих РНК являются разные гены, разбросанные по деконденсированным участкам хромосомных (точнее, хроматиновых) фибрилл.
Степень деконденсации может быть различной. Зоны полной деконденсации хромосом называются эухроматином. Он слабо воспринимает красители и практически не окрашивается. При неполном разрыхлении видны участки конденсированного хроматина, называемого гетерохроматином, который хорошо окрашивается и виден в ядре в виде базофильных глыбок. Эухроматин – рабочий активный хроматин, с участием которого в ядре в период интерфазы происходят процессы транскрипции и редупликации генетического материала. Гетерохроматин находится в состоянии покоя, выполняет функцию переноса генетического материала в дочерние клетки. Хроматин может равномерно распределять в ядре или располагаться отдельными глыбками.
Половой хроматин (тельце Барра) – дополнительная Х хромосома в соматических клетках женских особей, по которой можно определить принадлежность ткани или органа к полу (мужскому или женскому). Например, тельце Бара выявляется в большинстве нейтрофилов крови женщины.
Ядрышко представляет собой наиболее плотную структуру ядра округлой формы. Их может быть в ядре несколько, что зависит от метаболической активности клетки. Хорошо окрашивается основными красителями, так как содержит большое количество РНК и выполняет функцию синтеза рибосомных РНК и рибосом. В световом микроскопе ядрышко определяется как тонковолокнистая структура. В электронном микроскопе в нем выделяются два компонента: фибриллярный и гранулярный. Фибриллярный компонент располагается в центре ядрышка и представляет собой нитчатые структуры рибонуклеопротеидных тяжей предшественников рибосом. Гранулярный компонент занимает периферические части ядрышка и является созревающими субъединицами рибосом, окончательная сборка которых осуществляется в цитоплазме клетки. Ультраструктура ядрышка меняется в зависимости от уровня синтеза рРНК: чем он выше, тем больше гранул в нем образуется и наоборот.
В профазе митоза происходит исчезновения ядрышек и восстановление наблюдается в телофазе. Образование ядрышек связано с определенными участками хромосом – ядрышковыми организаторами, которые располагаются в зонах их вторичных перетяжек.
Ядеоный белковый матрикс. Представляет собой структурную сеть, состоящую из негистоновых белков, образующих основу, определяющую морфологию и метаболизм ядра. Он хорошо выявляется в интерфазных ядрах после растворения хроматина. Состоит из белковых фибрилл, располагающихся в периферическом слое нуклеоплазмы и образующих подстилающую пластинку – ламину. Кроме того, ядерный белковый матрикс образует внутриядерную сеть, к которой крепятся фибриллы хроматина. Его функция – поддержание общей формы ядра и организация пространственного расположения хромосом и их активности.
К клеточным мембранам относятся плазмолемма, кариолемма, мембраны митохондрий, эндоплазматической сети, аппарата Гольджи, лизосом и периксисом. Все биологические мембраны представляют собой тонкие слои (6–10 нм) липидов в комплексе с белками. На долю липидов в мембранах проходится 40 %, беков — 50–60 %, на многих мембранах обнаружены углеводы — до 5–10 %.
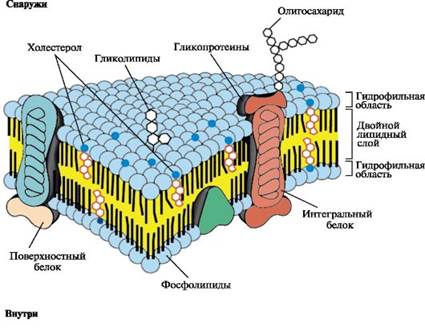
Составлипидоввразныхмембранахотличается. Основныелипидымембран:
—фосфолипиды (лецитин и цефалин) — состоят из гидрофильной головки (остаток фосфорной кислоты) и гидрофобного хвоста (2 остатка жирных кислот);
—сфингомиелины — состоят из сфингозина, холина, высших жирных кислот и фосфорной кислоты;
—холестерин (холестерол) — стероидный липид.
Основой любой мембраны являются фосфолипиды. Плазмолемма – внешняя клеточная мембрана — обогащена холестерином и сфингомиелином. Полярность липидов (наличие гидрофильной головки и гидрофобного хвоста) определяет их способность самопроизвольно образовывать двухслойные мембраны толщиной 5–7 нм.
Мембраны различаются набором белков. Многие белки также полярны, поэтому расположены в липидном слое в определенном порядке: неполярные части белков погружены в область хвостов фосфолипидов, гидрофильная часть белков находится рядом с головками фосфолипидов.
15
По расположению в билипидном слое все белки можно разделить на
3группы:
—интегральные — пронизывают оба фосфолипидных слоя;
—полуинтегральные — расположены в пределах одного слоя;
—периферические (примембранные) — не встроены в билипидный слой, только прилегают к нему.
Белки мембраны могут выполнять различные функции, поэтому они образуют функциональные группы: структурные белки, белки-рецепторы, белки-ферменты, белки-переносчики.
Углеводы мембран всегда связаны с молекулами липидов (гликолипиды) или белков (гликопротеиды).
Биологические мембраны обладают важнейшими свойствами:
—способность к самозамыканию — так как в мембране молекулы липидов и белков удерживаются гидрофобно-гидрофильными взаимодействиями, а не химическими связями, то при разрыве мембраны происходит
еесамопроизвольное восстановление;
—динамичность и текучесть мембран — в процессе жизнедеятель-
ности клетки от мембранных органелл отшнуровываются транспортные мембранные пузырьки с определенными веществами. Мембраны пузырьков легко встраиваются в мембраны других органелл и в плазмалемму. Происходит своеобразное перетекание мембран изнутри клетки наружу и в обратном порядке; молекулы в слоях мембраны способны к различным перемещениям. Так, свободно могут перемещаться липиды и белки в слое фосфолипидов — латеральное движение; известно спонтанное «флипфлоп» взаимодействие, когда молекулы фосфолипидов уходят из одного слоя в другой, белки-переносчики свободно перемещаются в толще билипидного слоя, осуществляя транспортную функцию; мембраны легко сжимаются и растягиваются при клеточных движениях;
—избирательная проницаемость — важнейшее свойство, регули-
рующее транспорт веществ. Для жирорастворимых веществ биологические мембраны не являются преградой. Эти вещества просто растворяются в билипидном слое. Для водорастворимых веществ мембрана является биологическим ситом.
Основные функции биологических мембран:
1.Барьерная функция. Биологические мембраны окружают все клетки снаружи, образуя плазмалемму.
2.Разграничительная функция. Мембраны разделяют клетку на участки — компартменты, различные по биохимическому составу, так образуются мембранные органеллы клетки: лабиринты эдоплазматической сети, стопки пузырьков аппарата Гольджи, мембранные пузырьки — лизосомы, пероксисомы, вакуоли. Двойная мембрана образует стенку митохондрий и ядра.
16
3.Функция синтеза. Ферменты мембран осуществляют важнейшие химические реакции, например, окислительное фосфорилирование при дыхании в митохондриях.
4.Рецепторная функция. С помощью рецепторов мембран распознаются различные физические или химические воздействия на клетку, происходит сортировка различные веществ, например, в аппарате Гольджи.
5.Транспортная функция мембран определяется ее важнейшим свойством — избирательной проницаемостью. Мембраны регулируют поступление веществ внутрь клетки и из нее.
Клеточная оболочка — это составная часть клетки, ее поверхностный аппарат. Поверхностный аппарат отграничивает клетку от внешней среды и состоит из 3-х компонентов:
1.Плазмолемма — липопротеиновый комплекс толщиной 10 нм (самая толстая из клеточных мембран), имеет вид трехслойной структуры — два электронно-плотных слоя разделены светлым слоем. Ее молекулярное строение описывается жидкостно-мозаичной моделью, согласно которой, мембрана состоит из липидного (фосфолипидного) бислоя, в который погружены и с которым связаны молекулы белков. В состав липидного слоя плазмолеммы входят холестерин и сфингомиелин.
2.Надмембранный комплекс представлен остатками углеводов, связанными с липидами и белками мембраны — это гликокаликс. Он придает поверхности клетки отрицательный заряд. Разветвленные углеводные молекулы являются рецепторами — воспринимающими молекулами. Рецепторы обеспечивают распознавание клеткой соседних клеток и межклеточного вещества, адгезивные взаимодействия с ними (адгезия — «прилипание»), способность воспринимать сигналы в виде биологически активных молекул — гормонов. В гликокаликсе энтероцитов кишечника могут адсорбироваться пищеварительные ферменты и происходить примембранное пищеварение.
3.Подмембранный комплекс — это тонкий слой цитоплазмы — кортикальный слой, в котором расположены ближе к мембране актиновые микрофиламенты цитоскелета, глубже — промежуточные филаменты и микротрубочки.
Кортикальный слой — специализированная периферическая часть цитоплазмы с ферментативными системами, связанными с трансмембранным транспортом и рецепторами. В кортикальном слое протекают важные для клетки синтетические и другие процессы, связанные с транспортом веществ в клетку и рецепцией биологически активных молекул. Актиновые микрофиламенты кортикального слоя связаны с белками мембраны и выполняют важную функцию — стабилизируют интегральные белки и обеспечивают их направленное перемещение. Благодаря сокращению сети микрофиламентов происходят изменения формы клетки, ее отдельных участков, формируются псевдоподии и выросты, что способствует процессам экзо- и эндоцитоза и перемещению клетки в пространстве.
Дата добавления: 2019-09-13; просмотров: 372; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!