Сигнальная система эритроцитов
Математические модели сигнальной системы эритроцитов
Эритроциты человека (красные клетки крови) — высокоспециализированные безъядерные клетки, которые участвуют в переносе кислорода из лёгких к тканям тела, а диоксида углерода в обратном направлении. Это самые многочисленные клетки крови. Их концентрация достигает 5х1012клеток/литр. Эритроциты ‑ элементы сложной многофункциональной системы, одна из функций которой поддержание в тканях концентрации кислорода и углекислого газа в заданном диапазоне значений в соответствии с запросами метаболизма. Это обеспечивается интенсивностью вентиляции легких, регулировкой работы сердца.
В эритроците обнаружены механо, потенциал и лиганд зависимые каналы, каналы, проводимость которых модулируется циклическими монофосфатами и т.д. Наличие таких элементов говорит о существовании сложной сигнальной системы (СС), управляющей параметрами эритроцитов. Сейчас известно, что эта СС управляет стационарными значениями агрегативности и пластичности эритроцитов.
При движении эритроцитов по кровяному руслу обратимо (т.е. триггерно) СС регулирует:
· агрегативность (AG) - способность формировать монетные столбики ,
· деформобильность PL(D) - способность менять форму под действием внешних сил
· проницаемость мембраны для кислорода и углекислого газа (PM)
Сигнальная система (СС) эритроцита, меняет стационарные значения AG, PL, PM. В свою очередь, эти параметры влияют на выполнение газотранспортной функции.
|
|
|
При механическом раздражении мембраны эритроцита, когда он входит в предкапиллярные артериолы и испытывает сжатие, срабатывает СС, которая переключает концентрацию кальция в цитоплазме эритроцита с  до
до  . Это изменение концентрации кальция приводит к тому, что уменьшается агрегативность, возрастает деформабильность и проницаемость мембраны для газов. Пока эритроцит идет по капилляру он остается в этом состоянии. При следующем механическом раздражении мембраны эритроцита, когда он выходит из капилляра в посткапиллярные венулы и начинает принимать исходную форму, срабатывает та же СС, которая переключает концентрацию кальция в исходное состояние с
. Это изменение концентрации кальция приводит к тому, что уменьшается агрегативность, возрастает деформабильность и проницаемость мембраны для газов. Пока эритроцит идет по капилляру он остается в этом состоянии. При следующем механическом раздражении мембраны эритроцита, когда он выходит из капилляра в посткапиллярные венулы и начинает принимать исходную форму, срабатывает та же СС, которая переключает концентрацию кальция в исходное состояние с 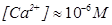 до и, как следствие при этом возрастает агрегативность и уменьшается деформабильность и проницаемость мембраны для газов.
до и, как следствие при этом возрастает агрегативность и уменьшается деформабильность и проницаемость мембраны для газов.
СС триггерно переключает параметры эритроцита с одного стационарного уровня на другой: в обменных капиллярах ‑ высокая PL и PM при низкой AG, в остальных отделах кровеносной системы: низкая PL и PM при высокой AG. За счет этого появляются дополнительные возможности оптимизации работы дыхательной системы, которые позволяют минимизировать затраты на продвижение крови по капиллярам, минимизировать газообмен со стенками сосудов и в тоже время позволяют обеспечить необходимую скорость газообмена в капиллярах.
|
|
|
На рисунке 2 представлена функциональная схема системы, управляющей AG, PL, PM эритроцитов. На этой схеме показано, что сигналы с рецепторов поступают на СС, затем анализируются, в результате вырабатываются управляющие воздействия, которые за счет фосфорилирования двух мембранных белков EF1, EF3 и белка цитоскелета EF2, изменяют AG, PL, PM. Причем под действием сигналов могут происходить как стационарные изменения AG, PL, PM, так и их триггерные переключения.

|
| Рисунок 2 – Функциональная схема СС эритроцита. М– механорецептор, Rs,i – рецепторы лигандов, управляющих параметрами эритроцитов, СС – сигнальная система, EF1, EF2, EF3 – белки, фосфорилирование которых приводит к изменению AG, PL, PM. |
Сигнальная система эритроцитов
На рисунке 3 представлена схема физико-химических процессов, происходящих в СС эритроцитов, управляющей фосфорилированием мембранных белков EF1, EF3 и белков цитоскелета EF2, которые в итоге определяют степень AG, PL и PM.

|
| Рисунок 3 - Схема сигнальной системы, определяющей уровень фосфорилирования определенных мембранных белков EF1, EF3 и белков цитоскелета EF2, которые определяют AG, PM и PL. Са, Na, Mg, K - ионы кальция, натрия, магния, калия, CaM – кальмодулин, cAMP – циклический аденозинмонофосфат, cGMP – циклический гуанидинмонофосфат, NOS – молекулярный модуль, формирующий концентрацию NO, ACII- молекулярный модуль, формирующий концентрацию cAMP, sGC – молекулярный модуль, формирующий концентрацию cGMP, Phos(EF1, EF2, EF3) – молекулярный модуль, фосфорилирующий белки, EF1, EF2, EF3, уровень фосфорилирования которых определяет AG, PL и PM, ICaA - каналы активного транспорта кальция, Icameс – механоактивируемые кальциевые каналы, Ica - потенциал активируемые кальциевые каналы, IKCaM – кальций кальмодулин зависимые калиевые каналы, IKV - потенциал зависимые калиевый каналы. AQP1 – аквапорины. |
Элементами СС являются физико-химические процессы, обусловленные работой определенных белков. Рассмотрим их по отдельности.
|
|
|
Показано, что у эритроцитов есть мембранные белки AQP1 ‑ аквапорины тип 1 – высокоселективные водные каналы. Через эти каналы может проходить вода, кислород и углекислый газ, и они не проницаемы для заряженных молекул. При образовании комплекса AQP 1 с циклическими монофосфатами, прежде всего с cGMP, AQP 1 становятся проницаемыми для катионов. Показано, что фосфорилирование AQP1 протеинкиназами PKA , PKC и PKG приводит к увеличению проницаемости этого канала для воды, для кислорода и углекислого газа.
|
|
|
В мембране эритроцитов есть несколько типов кальциевых каналов: механоактивируемые Icameс, потенциалозависимые Ica и кальций кальмодулин зависимый каналы активного транспорта кальция ICaA. В мембране эритроцитов есть кальций зависимые калиевые каналы IKCaM, потенциал зависимые калиевые каналы IKV. В эритроцитах есть белок кальмодулин CaM и есть процессы, обеспечивающие изменения концентрации циклических монофосфатов (cAMP, cGMP).
Ø Проанализируем работу этой сигнальной системы.
Моделирование
На основе схемы, представленной на рисунке 3, была построена математическая модель, состоящая из двух обыкновенных дифференциальных уравнений:
для  ‑ скорости изменения концентрации кальция в цитоплазме эритроцита, и для
‑ скорости изменения концентрации кальция в цитоплазме эритроцита, и для  ‑ скорости изменения трансмембранной разности потенциалов.
‑ скорости изменения трансмембранной разности потенциалов.
 (1)
(1)
Найдены такие значения параметров, при которых система работает как триггер, с двумя устойчивыми значениями стационарных концентраций кальция: одна при » 10-7М, другая при » 10-6М. При этом трансмембранная разности потенциалов около -50 мВ. Переключения из одного стационарного состояния в другое и обратно происходит примерно одинаковыми кальциевыми импульсами.
Анализ математической модели
Уравнения (1) при варьировании значений параметров в широком диапазоне значений, дают три принципиально разных типа фазовых портретов:
1) одна особая точка (устойчивый узел) при значении концентрации кальция  ;
;
2) три особых точки (два устойчивых узла). Первая устойчивая особая точка при значении концентрации кальция  вторая устойчивая особая точка при
вторая устойчивая особая точка при 
3) одна особая точка (устойчивый узел) при значении концентрации кальция  .
.



Рисунок 4 – Фазовые портреты системы (1) при трех разных значениях параметров. A) – одна особая точка a – устойчивый узел в области малых значений концентрации кальция. B) - три особые точки a , b , c. Особые точки a , c – устойчивые узлы. С) – одна особая точка c – устойчивый узел в области средних физиологических значений концентрации кальция.
В частности, когда есть три особых точки a , b , c переключение из особой точки a в c и обратно может происходить прямоугольным импульсом кальциевого тока определенной амплитуды A и длительности T. Амплитуда импульса изменяет количество особых точек. При Acon ≥ 0.01884 у системы (1) есть только одна особая точка с. При меньших значениях Acon три особых точки.
В зависимости от амплитуды А и длительности импульса при раздражении клетки наблюдается изменение концентрации кальция (ca) во времени (t).
Найдены значения параметров, при которых система (1) имеет три особые точки a , b , c, (см. рис.4В) расположенные по оси потенциалов в области -2 (-50 mV), по оси концентраций кальция a (ca » 0.1), c (ca » 1). Найдены значения параметров, при которых система триггерно переключается из a в c и обратно. Эти значения параметров СС эритроцита можно отнести к одному из вариантов нормы. В этом случае переключение параметров эритроцитов будет способствовать более оптимальному выполнению газотранспортной функции.
Ø При этом необходимо учитывать, что все параметры реальных эритроцитов (плотность каналов, общие концентрации ферментов и т.д.) непрерывно изменяются. Поэтому, реальные эритроциты будут иметь разброс всех параметров, который приведет к разным значениям стационарных концентраций кальция.
При различных патологиях, при старении, и при действии физиологически активных соединений будут происходить изменения в значениях параметров СС. Могут возникать ситуации, когда есть только одна особая точка a или c. В этом случае в ответ на раздражение кальциевым импульсом система будет отвечать только импульсным изменением ее характеристик.
Показано, что по мере старения эритроцитов в них увеличивается концентрация кальция. Если у молодых эритроцитов она составляет в среднем 0.62•10-7M, то у старых около 2•10-7M, эти значения в области стационарной особой точки a (рис. 4b). О каких-то триггерных переключениях в этом случае говорить нельзя. Скорее всего, это указывает на то, что по мере старения эритроцитов система кальциевого гомеостаза деградирует.
Сейчас показано, что на AG эритроцитов влияют как состояние определенных мембранных белков, так и концентрации определенных белков плазмы крови, покрывающих поверхность клетки. На AG будет также оказывать влияние разброс эритроцитов по состояниям с разной степенью фосфорилирования белков. Эритроциты, находящиеся в особой точке c в сосудах будут способствовать обрыванию роста монетных столбиков, поскольку в этом состоянии агрегативность низкая. Чем больше эритроцитов находится в состоянии c , когда они идут по крупным сосудам, тем будет короче средняя длина монетных столбиков. И когда все эритроциты в c, то монетные столбики будут минимальными. В этом случае будет и минимальная скорость осаждения эритроцитов.
Таким образом, можно заключить, что если у эритроцита есть СС, подобная той, которая указана на рис. 3, то у нее существуют такие значения параметров, при которых AG, PL и PM переключаются из одного стационарного состояния в другое триггерно с примерно одинаковыми порогами переключения из a в с и обратно.
Дата добавления: 2019-02-22; просмотров: 302; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!