Низхідні провідникові шляхи, через які здійснюються впливи центрів головного мозку на рухові центри спинного мозку
МІНІСТЕРСТВО ОХОРОНИ ЗДОРОВ’Я УКРАЇНИ
КИЇВСЬКИЙ МЕДИЧНИЙ УНІВЕРСИТЕТ УАНМ
| “Затверджено” на методичній нараді кафедри нормальної фізіології Завідувач кафедри професор____________ З.Д. Скрипнюк “______“ серпня 2013 р. |
МЕТОДИЧНІ ВКАЗІВКИ
ДЛЯ САМОСТІЙНОЇ РОБОТИ СТУДЕНТІВ ПРИ ПІДГОТОВЦІ ДО ПРАКТИЧНОГО ЗАНЯТТЯ
| Навчальна дисципліна | “ФІЗІОЛОГІЯ” |
| Модуль № 1 | “ЗАГАЛЬНА ФІЗІОЛОГІЯ” |
| Змістовий модуль №4 | Роль центральної нервової системи (ЦНС) у регуляції рухових функцій. |
| Тема практичного заняття №8 | Дослідження ролі спинного мозку в регуляції рухових функцій організму. |
| Курс | 2-й |
| Факультет | Медичний |
Київ – 20 13
1.Актуальність теми:
Спинний мозок у ЦНС є першим рівнем регуляції фізіологічних функцій, і зокрема, рухових функцій. Саме тут розташовані нервові центри, на які здійснюють свій влив нервові центри головного мозку, передаючи інформацію низхідними провідними шляхами і забезпечуючи пристосувальні реакції організму. В той же час нервові центри головного отримують інформацію висхідними шляхами про стан регульованих параметрів, що забезпечує корекцію параметрів пристосувальної реакції.
У разі ураження провідних шляхів спинного мозку, що забезпечують його зв’язки з центрами спинного мозку характер спінальних рефлекторних реакцій змінюється, або вони зовсім не здійснюються при пошкодженні спінальних центрів
2.Навчальні цілі:
Ø Робити висновки про стан рухових функцій організму - м’язового тонусу, пози, локомоцій, рухових рефлексів що мають місце після поперечного перерізу на різних рівнях ЦНС та пошкодження рухових структур
Ø Аналізувати регульовані параметри та механізми активації рецепторів як слідкуючих пристроїв при здійсненні рухових рефлексів.
Ø Робити висновки про стан рухових рефлексів, які замикаються на різних рівнях ЦНС, та будову їх рефлекторних дуг.
Ø Робити висновки про роль провідних шляхів ЦНС у забезпеченні сенсорних і рухових функцій організму.
Базовий рівень підготовки
Студенти мають навики з попередніх навчальних дисциплін:
| Назви попередніх дисциплін | Отримані навики |
| Анатомія людини | Аналізувати інформацію про будову тіла людини, системи, що його складають, органи і тканини |
| Гістологія, цитологія, ембріологія | Інтерпретувати мікроскопічну та субмікроскопічну структуру клітин |
| Медична біологія | Пояснювати закономірності проявів життєдіяльності людського організму на молекулярно-біологічному та клітинному рівнях. |
Завдання для самостійної праці під час підготовки до практичного заняття
Перелік основних термінів, параметрів, характеристик, які повинен засвоїти студент при підготовці до практичного заняття
| Термін | Визначення |
| Спінальний шок | Це тимчасова відсутність спінальних рефлекторних реакцій, що виникає нижче місця зв’язків спинного мозку з головним мозком. |
| Тонічні рухові рефлекси | Забезпечують підтримання м’язового тонусу. |
| Фазичні рухові рефлекси | Забезпечують переміщення кінематичних пар або кінематичних ланцюгів завдяки скороченню м’язів. |
| Кінематична пара | Це дві кістки, що з’єднані суглобом. |
| Кінематичний ланцюг | Це сукупність декількох кінематичних пар (наприклад, кінцівки) |
| Міотатичні рефлекси або рефлекси на розтягнення. | Це рефлекси, які виникають при розтягненні рецепторів м’язів - м’язових веретен. |
| Нейромоторна одиниця | Це група м’язових волокон, що іннервується одним мотонейроном. |
Теоретичні питання
1) Поняття про позу тіла й умови її забезпечення. Установчі позні та рухові рефлекси організму.
2) Види сенсорної інформації, що надходять у спинний мозок від рецепторів організму.
3) Рефлекторна діяльність спинного мозку, види спінальних рефлексів. Тонічні і фазичні рухові рефлекси спинного мозку, їх фізіологічне значення і будова рефлекторних дуг. Ефективність тонічних рухових рефлексів спинного мозку при автономній його діяльності.
4) Залежність спінальних рефлексів від діяльності центрів головного мозку. Спінальний шок.
5) Медіальна та латеральна низхідні системи регуляції рухових функцій організму. Характер впливів кожного із шляхів цих систем на спінальні а-мотонейронирізного функціонального значення. Способи зв’язку волокон низхідних шляхів із спіральними а-мотонейронами.
Практичні роботи
1) Дослідження спінального шоку у жаби.
2) Дослідження м’язового тонусу спінальної жаби.
3) Дослідження пози спіральної жаби.
4) Дослідження фазичних шкірно-м’язових рухових рефлексів спіральної жаби.
5) Дослідження сухожильних рефлексів у людини.
Зміст теми
Регуляція рухових функцій за участю центральної нервової системи (ЦНС) - це регуляція:
тонусу скелетних м’язів,
пози тіла,
переміщення частин тіла відносно тулуба,
переміщення тіла в просторі - локомоцій.
Різні відділи ЦНС здійснюють регуляцію рухових функцій, впливаючи на нейромоторні одиниці.
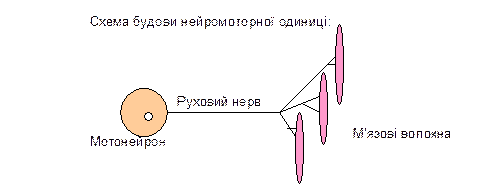
Кількість м’язових волокон у нейромоторних одиницях різна - від декількох одиниць до десятків, сотні, тисяч, що пов’язано з регуляцією сили скорочення м’язів та їх функцією. М’язові волокна моторних одиниць належать до
а) швидких
б) повільних.
Швидкі м’язові волокна
- мають тривалість одиночного скорочення близько 20-50 мс,
- можуть скорочуватися в анаеробних умовах ,
- розвивають значну силу,
- швидко втомлюються.
Повільні м’язові волокна
- мають тривалість одиночного скорочення 100-200 мс,
- скорочуються в аеробних умовах,
- розвивають меншу силу,
- довго не втомлюються.
Скелетні м’язи людини за будовою змішані - до їх складу входять як швидкі, так і повільні м’язові волокна.
Мотонейрони розташовані в передніх рогах спинного мозку і належать до двох типів:
1.Малі мотонейрони: іннервують малу кількість м’язових волокон; мають низькі пороги і активуються найпершими; генерують ПД з невеликою частотою, що призводить до меншої сили скорочення м’язових волокон.
2. Великі мотонейрони: іннервують значну кількість м’язових волокон; мають високі пороги і активуються пізніше; генерують ПД з більшою частотою, що призводить до більшої сили скорочення м’язових волокон.

А.Сенсорна функція спинного мозку: центральний аналіз інформації від рецепторів
Спинний мозок отримує інформацію від пропріорецепторів про стан опорно-рухової системи та екстерорецепторів шкіри - про контакт з зовнішнім середовищем і забезпечує здійснення спінальних рухових рефлексів.
До пропріорецепторів належать:
а. М’язові веретена - розташовані паралельно м’язовим волокнам і передають інформацію по аферентних нервових волокнах групи Аa про зміни довжини м’язів (статичні зміни) та швидкість їх розтягнення (динамічні зміни).
б. Сухожильні рецептори Гольджі - розташовані в сухожиллях, мають високі пороги і передають інформацію по аферентних нервових волокнах групи Аb про розтягнення сухожиль при скороченні м’язів.
в. Тільця Пачіні - розташовані в м’язах, передають інформацію по аферентних нервових волокнах групи В про вібрацію, подібні до них пропріорецептори розташовані в суглобових сумках і зв’язках і передають інформацію про зміни руху в суглобах (швидкість руху та величину кута) .
г. Вільні нервові закінчення - розташовані в м’язах, вони високо порогові, належать до ноціцепторів - рецепторів пошкодження, передають інформацію по аферентних нервових волокнах групи В, С.
М’язові веретена, їх будова. Їх кількість у різних скелетних м’язах різна. Основними структурними елементами м’язового волокна є інтрафузальні м’язові волокна, нервові волокна і капсула.
|
|
|
|
|
|
|
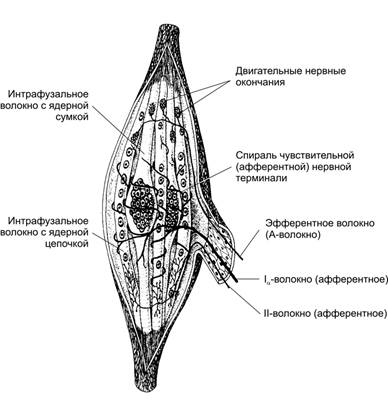
Схема будови м’язового веретена. До веретена підходять аферентні та еферентні нервові волокна. Спіралевидні (первинні) нервові волокна утворенні нервовими закінченнями аферентних волокон групи Аа і відходять від центральної ділянки інрафузальних м’язових волокон (МВ), вони передають інформацію про швидкість та розтягнення та довжину м’язів. Ближче до кінців інтрафузальних м’язових волокон розташовані вторинні нервові закінчення, які переважно передають інформацію від інтрафузальних м’язових з ядерним ланцюжком. Еферентні Аg-волокна утворюють нервово-м’язові синапси з інтрафузальними МВ в їх дистальній частині.
М’язове веретено має від 1 до 10 коротких інтрафузальних м’язових волокон: середній частині вони мають скупчення ядер (волокна з ядерною сумкою), або ядра розташовані ланцюжком (волокна з ядерним ланцюжком).
Капсула. Комплекс інтрафузальних МВ с нервовими терміналями оточений капсулою, зовнішній шар якої є похідним периневрія, а внутрішні – це аналоги ендоневрія.
При розтягненні м’язів розтягуються інтрафузальні м’язові волокна і передають інформацію від спіралевидних рецепторів по аферентним нервовим волокнам до а-мотонейронів спинного мозку , активація а-мотонейронів призводить до скорочення екстрафузальних м’язових волокон - зменшення їх довжини.
б. гама - мотонейрони отримують інформацію від різних структур головного мозку і передають нервові імпульси до інтрафузальних м’язових волокон, які скорочуються і розтягують їх ядерну сумку , забезпечуючи тим самим можливість передачі інформації під час скорочення м’язів до а-мотонейронів та інших відділ ЦНС.
в. а-мотонейрони і гама-мотонейрони активуються одночасно сигналами від рухових структур головного мозку (коактивація), що дозволяє підтримувати розтягненими м’язові веретена під час скорочення м’язів і передавати весь час інформацію про динамічні і статичні зміни м’язів .
При розтягненні м’язів розтягуються інтрафузальні м’язові волокна і передають інформацію від спіралевидних рецепторів по аферентним нервовим волокнам до а-мотонейронів спинного мозку , активація а-мотонейронів призводить до скорочення екстрафузальних м’язових волокон - зменшення їх довжини.
б. гама - мотонейрони отримують інформацію від різних структур головного мозку і передають нервові імпульси до інтрафузальних м’язових волокон, які скорочуються і розтягують їх ядерну сумку , забезпечуючи тим самим можливість передачі інформації під час скорочення м’язів до а-мотонейронів та інших відділ ЦНС.
в. а-мотонейрони і гама-мотонейрони активуються одночасно сигналами від рухових структур головного мозку (коактивація), що дозволяє підтримувати розтягненими м’язові веретена під час скорочення м’язів і передавати весь час інформацію про динамічні і статичні зміни м’язів .
Сухожильні рецептори Гольджи розташовані у сухожиллях на межі з м’язом. Рецептор має веретеноподібну будову і оточений капсулою. Терміналі аферентних нервових мієлінових волокон розташовані серед пучків колагенових волокон, що розташовані у заповненому рідиною просторі. Рецептори активуються при розтягненні сухожилля м’язом, який скорочується, що призводить до рефлекторного розслаблення цього м’язу.
|
|
|
|
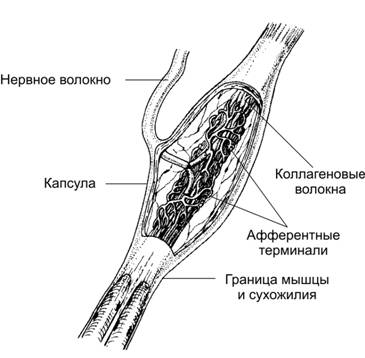
Схема структури сухожильного рецептору Гольджи.
В. Рефлекторна функція спинного мозку.
СПІНАЛЬНІ РУХОВІ РЕФЛЕКСИ
В центрах спинного мозку замикаються рефлекторні дуги рухових рефлексів:
а) тонічних, що лежать в основі регуляції тонусу м’язів;
б) фазичних, що забезпечують переміщення частин тіла відносно тулуба.
Спінальні рефлекси є сегментарними або міжсегментарними.
1) Рефлекси розтягнення з м’язових веретен – міотатичні:
- виникають при розтягненні м’язових веретен внаслідок розтягнення м’язів;
- інформація від м’язових веретен (первинних спіралеподібних нервових закінчень) по аферентним нервовим волокнам передається до а-мотонейронів спинного мозку, які активуються;
- активація а-мотонейронів призводить до передачі інформації до скелетних м’язів по еферентним нервовим волокнам, наслідком цього є їх скорочення;
- альфа-мотонейрони м’язів-антагоністів гальмуються;
- рефлекторна дуга міотатичних рефлексів, що виникають при розтягненні м’язових веретен, є моносинаптичною;
- при відсутності низхідних активуючих впливів рухових центрів головного мозку на а-мотонейрони та гама-мотонейрони міотатичні рефлекси підтримують м’язовий тонус, але він є ослабленим.
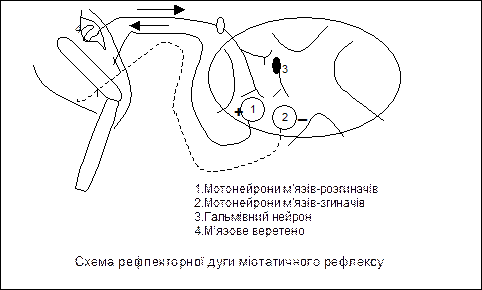
2) Рефлекси розтягнення з рецепторів сухожиль:
- виникають при розтягненні сухожиль м’язом, який скорочується;
- інформація від рецепторів сухожиль Гольджі передається по аферентним нервовим волокнам до гальмівних нейронів спинного мозку, які викликають постсинаптичне гальмування а-мотонейронів м’язу, що скорочувався, наслідком цього є його розслаблення. Рефлекторна дуга рефлексу є полісинаптичною.
|
|
|
|
|
|
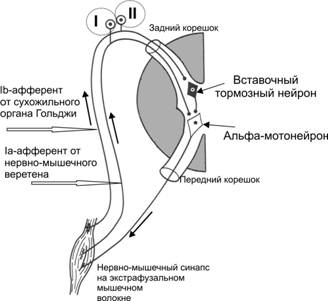
Схеми рефлекторних дуг: 1) рефлексу розтягнення (І) з м’язових веретен (що призводить до скорочення того ж м’язу ; 2) гальмівного рефлексу розтягнення з рецепторів сухожиль Гольджі (II), яка має гальмівний нейрон, що призводить до гальмування а-мотонейрону і розслаблення м’язу, який скорочувався.
3) Флексорні шкірно-м’язові рефлекси:
- виникають при подразненні рецепторів шкіри
інформація передається по аферентним волокнам до альфа-мотонейронів спинного мозку, які активуються;
- від альфа-мотонейронів нервові імпульси передаються по еферентним нервовим волокнам до м’язів-флексорів, які скорочуються, наслідком чого є згинання кінцівки та віддалення її від подразника;
- рефлекторні дуги шкірно-м’язових рефлексів є полісинаптичними.

С.Провідникова функція спинного мозку.
Провідні шляхи утворені ланцюгами нейронів, що з’єднані між собою синапсами й забезпечують передачу інформації від нейрону до нейрону, від ядра до ядра.
Висхідні провідникові шляхи
Утворені центральними відростками аферентних нейронів спинномозкових вузлів:
ò Тонкий пучок (Голля, fasciculus gracilis) проходить у складі заднього канатика і закінчується в тонкому ядрі довгастого мозку; провідний шлях пропріоцептивної і тактильної чутливості.
ò Клиноподібний пучок (Бурдаха, fasciculus cuneatus) проходить у задньому канатику і закінчується в клиноподібному ядрі довгастого мозку; провідний шлях пропріоцептивної й тактильної чутливості.
Утворені аксонами вставних нейронів:
ò Спіноталамічний шлях вентральний (tractus spinothalamicus ventralis) — проекційный афферентний шлях, що проходить у передньому канатику. Периферичні відростки перших нейронов, розташовані у спинномозкових вузлах, проводять тактильні і пресорні відчуття від механорецепторів шкіри. Центральні відростки цих нейронів входять через задні корінці у задні канатики, де піднімаються на 2–15 сегментів і утворюють синапси з нейронами задніх рогів. Аксони цих нейронів переходять на протилежну сторону в складі передньої спайки й проходять далі в передній периферичній зоні передньобокових канатиків. Звідси волокна піднімаються до задньолатерального вентрального ядра таламуса разом с латеральним спіноталамічним шляхом.
ò Спіноталамічний шлях латеральний (tractus spinothalamicus lateralis) — проекційний аферентний шлях, що проходить у боковому канатику. Периферичними рецепторами є вільні нервові закінчення шкіри. Центральні відростки псевдоуніполярних нейронів спинномозкових вузлів входять у спинний мозок через латеральні відділи задніх корінців і, піднявшись у спинному мозку на 1–2 сегменти, утворюють синапси з нейронами задніх рогів. Аксони цих нейронів утворюють латеральний спіноталамічний шлях. Вони йдуть через передню спайку на протилежну сторону й піднімаються в латеральних відділах бокових канатиків. Спіноталамичні шляхи проходять через стовбур мозку й закінчуються в вентро-латеральних ядрах таламуса. Це головний шлях проведення больової й температурної чутливості.
|
|
|
|

|

|
|
|
|
|
|
|
|
|
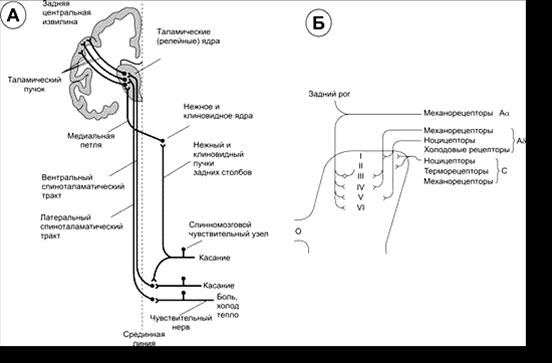
ò Спіномозочковий шлях передній (tractus spinocerebellaris anterior) — проекційний аферентний пропріоцептивний шлях мозочка, що проходить у латеральному канатику; входить після перехресту у верхню ніжку мозочка протилежної сторони й закінчується в корі мозочка.
ò Спіномозочковий шлях задній (пучок Флексига, tractus spinocerebellaris posterior) — проекційний аферентний пропріоцептивний шлях мозочка, що проходить у боковому канатику спинного мозку й нижній ніжці мозочка; закінчується в корі мозочка тієї ж сторони. Цим шляхом до мозочка йде інформація від усіх пропріорецепторів, що необхідно для координації довільних рухів.
ò Спінооливний шлях (tractus spino-olivaris) — пучок висхідних волокон переднього канатика, що закінчується у додаткових ядрах оливи.
ò Спінопокришечний (спінотектальний) шлях (tractus spinotectalis) — проекційний висхідний шлях загальної чутливості, що проходить у передньому канатику, стовбурі головного мозку й закінчується в нижніх и верхніх горбиках середнього мозку протилежної сторони.
ò Спіноретикулярний шлях (tractus spinoreticularis) — пучок висхідних волокон бокового канатика, що закінчується в ретикулярній формації довгастого мозку, моста и середнього мозку. Шлях надходження аферентної інформації соматичних і вісцеральних рефлексів.
Низхідні провідникові шляхи, через які здійснюються впливи центрів головного мозку на рухові центри спинного мозку
- Пірамідні шляхи (кортико-спінальні, кортико-бульбарні) проходять через довгастий мозок на рівні перехресту пірамід і несуть інформацію від моторної зони кори .

| |


|

|
|
|
|
|


|
|
|
|
| |
|
|
|
|
|
|
|
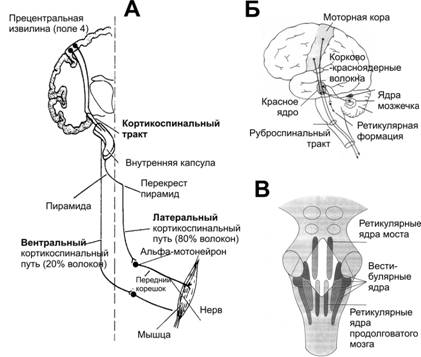
Схема низхідних рухових шляхів мозку: А. Кортикоспінальний (пірамідний) тракт. Б. Кортикоруброспінальний шлях. В. Розташування ретикулярних і вестибулярних ядер у стовбурі мозку.
Всі інші рухові шляхи починаються від структур стовбуру мозку:
- а.Рубро-спінальний тракт
- передає інформацію від червоних ядер до латеральних інтернейронів спинного мозку;
- стимуляція червоних ядер призводить до активації мотонейронів флексорів і гальмування мотонейронів екстензорів.
- б.Понтінно-ретикуло-спінальний тракт
- починається від ядер мосту і передає інформацію до вентромедіальних відділів спинного мозку;
- стимуляція ядер мосту призводить до активації мотонейронів як флексорів та екстензорів з переважним впливом на активацію мотонейронів екстензорів.
- в.Медулярний ретикуло-спінальний тракт
- починається від ретикулярної формації стовбуру мозку і передає інформацію до інтернейронів спинного мозку;
- стимуляція має загальний гальмівний вплив, переважно на мотонейрони екстензорів.
- г.Латеральний вестибуло-спінальний тракт
- починається від ядер Дейтерса і передає інформацію на інтернейрони і мотонейрони з тієї ж сторони;
- стимуляція ядер Дейтерса призводить до активації мотонейронів екстензорів і гальмування мотонейронів флексорів.
- д.Текто-спінальний тракт
- починається від верхніх двогорбиків і передає інформацію до мотонейронів шийного відділу спинного мозку
- забезпечує регуляцію функції шийних м’язів.
Дата добавления: 2019-02-12; просмотров: 136; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!