Регуляція статевої функції
Органи розмноження сільськогосподарських тварин перебувають під постійним впливом нервової і гуморальної (ендокринної) систем організму, взаємодія яких і обу-мовлює прояви і повноцінність у них статевої функції. Нервова регуляція реалізуєть-ся за загальними принципами рефлекторної діяльності – через рецептори, аферентні шляхи, статеві центри та ефекторні провідники. Вся інформація врешті сумується, аналізується і реалізується через кору головного мозку.
Загальна регуляція статевої функції у тварин здійснюється центральною нерво-вою системою шляхом складних нейросекреторних процесів, що перебігають суворо ритмічно у гіпоталамо-гіпофізарній системі з виділенням гонадотропін-рилізінг фак-тора (гонадотропін-рилізінг гормону, Гн–РГ) у гіпоталамусі, гонадотропних гормонів у гіпофізі та статевих гормонів у гонадах.
Крім згаданих гормонів у регуляції статевої функції беруть також участь гормони наднирників, щитовидної залози, тимусу, проте вони впливають на ці процеси непря-мим шляхом. Останнім часом увагу дослідників привертає епіфіз – нейроендокринна залоза, що впливає на статеве дозрівання ссавців, але деталі цього процесу недостат-ньо вивчені. Відомо лише, що через нього реалізуються світлові впливи.
“Ініціатором” статевої циклічності у самок є гіпоталамус.
Нарешті, істотна роль належить також локальним клітинним та міжклітинним факторам взаємодії, наявним у яєчниках (зокрема у теці та гранульозі фолікула) та криптах ендометрію. Найбільш вивченими серед них є простагландини типу F2α.
Гіпоталамус (основна частина проміжного мозку) вважають вищим статевим центром (біологічним годинником). Це своєрідний нейроендокринний орган, що за-безпечує єдність нервових та ендокринних процесів.
У гіпоталамусі розрізняють три області: передню, що містить супраоптичні та па-равентрикулярні ядра; середню (область сірого горба) з вентро-медіальним та аркуат-ним ядром, та задню область з мамілярними ядрами. Центри, що регулюють гонадо-тропну функцію гіпофіза, в основному розміщені у передньому гіпоталамусі. Так, су-праоптичні ядра переднього гіпоталамусу синтезують вазопресин (антидіуретичний гормон, АДГ), який, поряд з посиленням зворотного всмоктування води у ниркових каналиках, зниженням діурезу, регуляцією мінерального обміну, підвищенням арте-ріального тиску, викликає також скорочення матки та кровоносних судин. Виділяєть-ся гормон рефлекторно при больових та інших стресових станах. Ядра середнього гіпоталамуса регулюють тонічне (постійне) виділення гіпофізом гонадотропінів, тоді як ядра преоптичної області забезпечують циклічне їх виділення.
Розділ 2
У паравентрикулярних ядрах цієї ж області виробляється переважно окситоцин, що виділяється рефлекторно при ссанні, доїнні, спаровуванні. Він викликає скорочен-ня матки, піхви, міоепітеліальних клітин молочної залози, а у самців – сім’япроводу.
Важлива роль у кірково-гіпофізарній регуляції належить медіаторам та тим фер-ментним системам, які беруть участь у обміні гормонів. Проміжними ланками у ме-ханізмі реакцій відповіді на дію гормонів є простагландини. Нині відомо 20 проста-гландинів, що належать до 4 груп (А, В, E, F), що діють по-різному. З них проста-гландини Е та F беруть участь у регуляції статевої функції. Вони стимулюють синтез пролактину, прогестерону і інших сполук, зменшують інкрецію лютропіну, беруть участь у передачі нервових імпульсів, впливають на роботу серця, тонус судин. ПГ-Ф збільшує, а ПГ-Е зменшує скоротливість матки. ПГ-Ф та ПГ-Е регулюють акт родів. Простагландини F2α виробляються в кінці статевого циклу у ендометрії і викликають розсмоктування жовтого тіла.
Синтез та виділення Гн–РФ визначаються співвідношенням дофаміну та серо-тоніну.
Нервові клітини гіпоталамусу продукують біологічно активні речовини пептичної природи – нейрогормони (рилізінг-гормони). Сприймаючи інформацію про стан орга-нізму та довкілля, гіпоталамус аналізує її, інтегрує і перетворює у рилізінг-гормони.
Нервові клітини гіпоталамусу виробляють біля 10 рилізінг-гормонів, що через гіпофіз контролюють секрецію гормонів у гонадах та інших залозах внутрішньої се-креції. Одні з них – ліберини – стимулюють вивільнення з аденогіпофізу у кров від-повідних гормонів (фоліберин, пролактоліберин, тиреоліберин і т. ін), інші – статини (пролактостатин, соматостатин, меланостатин) – гальмують їх виділення. Проте, го-ловне значення у регуляції статевого циклу має гонадоліберин, що володіє активністю фоліберину та люліберину. Він регулює виділення гіпофізом цих гонадотропінів.
Продукований у гіпоталамусі Гн–РГ транспортується через ворітну систему до аденогіпофіза, вивільнюючи синтезовані тут гонадотропіни. Останні виконують функцію медіаторів, які передають імпульси з гіпоталамуса на яєчники. Виділення Гн–РГ у гіпоталамусі хоча й регулюється центральною нервовою системою, проте перебуває у тісному взаємозв’язку з гормональною активністю гонад. Секреція Гн–РГ має переривчастий (імпульсний) характер, що обумовлює імпульсну секрецію фолі-тропіну та лютропіну. Під час фолікулінової фази циклу відбувається різке зростання частоти імпульсної секреції Гн–РГ.
Гіпофіз. У 1912 р. Ашгейм встановив, що екстирпація гіпофіза у собак викликає атрофію яєчників, сім’яників та зникнення вторинних статевих ознак. Опісля Сміт і Цондек, підсаджуючи тканину гіпофіза оперованим тваринам, викликали у них по-яву тічки та охоти. Згодом вдалося виділити з гіпофіза білкові речовини, що володіли гонадотропною дією: одна з них стимулювала розвиток фолікулів і була названа фолі-кулостимулюючим гормоном (ФСГ, нова назва фолітропін), інша дістала назву люте-їнізуючого гормону (ЛГ, нова назва лютропін), тому що вона викликала лютеїнізацію дозрілих фолікулів.
Фізіологічні особливості статевих органів свійських тварин
Гонадотропні гормони (гонадотропіни) утворюються у невеликих кількостях у передній долі гіпофіза постійно, протягом усього життя тварин. У період статевого дозрівання і під час окремих стадій статевого циклу секреція їх значно зростає. Вони обумовлюють розвиток статевих органів, підтримують їх функцію і стимулюють біо-синтез статевих гормонів. У подібному випадку говорять про циклічність функції гі-пофізу. Під час вагітності гонадотропні гормони виробляються у гіпофізі у значно більших кількостях (у 8–10 разів) і в дуже великих кількостях у плаценті, тому їх ділять на гіпофізарні та позагіпофізарні (плацентрані) гонадотропіни.
Гіпофіз має подвійну регуляцію: симпатична система гальмує виділення гонадо-тропних гормонів, а парасимпатична – підсилює їх виділення. Характер регуляції змі-нюється по фазах циклу: естрогени стимулюють парасимпатичну нервову систему, а прогестерон – симпатичну.
Під час тічки й охоти фолітропін і лютропін накопичуються у крові різних тварин в неоднакових співвідношеннях, що обумовлює різну тривалість у них цих стадій. Так, у гіпофізі корів, овець і свиней переважає виділення лютропіну над фолітропі-ном, тоді як у кобил і кролиць – навпаки, переважає фолітропін.
Фолітропін та лютропін проявляють свою дію в організмі комплексно. Співвід-ношення їх у гіпофізі визначає особливості статевого циклу у тварин. Так, високий вміст фолітропіну та низький лютропіну у кобил обумовлює збільшення тривалості охоти, а овуляція у них наступає задовго до закінчення охоти. У корів, навпаки, вміст фолітропіну низький, а лютропіну високий, тому охота у них коротша, іноді навіть статевий цикл буває алібідним.
З функціональної точки зору гонадотропні гормони подібні між собою, але вони володіють різною фармакологічною і фізіологічною дією.
У самців під впливом високих концентрацій тестостерону втрачається циклічний тип функціонування гіпоталамуса.
Фолікулостимулюючийгормончифолітропін (ФСГ, раніше його називали про-лан А) є альбуміном з вуглеводним радикалом. Він стимулює овогенез, ріст і розвиток вторинних фолікулів до антральної стадії. Проте, під впливом тільки фолітропіну фо-лікули не досягають стадії повного росту і секреторної активності, а лише готуються до подальшого впливу лютропіну.
У самців фолітропін стимулює розвиток покручених каналиків сім’яників таспер-міогенез.
Лютеїнізуючийгормончилютропін (ЛГ, стара назва пролан Б) є глюкопротеї-дом. Він прискорює дозрівання фолікулів, викликає лютеїнізацію фолікулярних клі-тин, овуляцію і утворення жовтого тіла, його дія буває чітко вираженою лише після попередньої дії фолітропіну.
У самців лютропін стимулює секреторну діяльність інтерстиціальних клітин сім’яників (клітин Лейдіга).
Механізм дії у яєчнику фолітропіну та лютропіну можна уявити собі таким чи-ном: вони захоплюються тут гормонозалежними клітинами – мішенями, і розміщені
Розділ 2
на їх поверхні циторецептори утворюють з молекулою гормону комплекс, що про-никає у ядро клітини, взаємодіє з її геномом, викликає специфічний ефект, після чого руйнується і витісняється з клітини.
Концентрація фолітропіну та лютропіну у крові корів та свиней змінюється від-повідно до фаз статевого циклу: з 15–17-го дня циклу різко зростає вміст фолітропіну. За день до охоти і під час неї зростає концентрація лютропіну; на другий день циклу концентрація гонадотропінів різко знижується, а з 11–13-го дня спостерігається дру-гий підйом концентрації гонадотропінів (тривалістю для фолітропіну 1–2 дні, а для лютропіну – 1–3 дні), що зв’язане з початком регресії жовтого тіла і розвитком нових фолікулів.
Лютеотропнийгормон (ЛТГ або пролактин, лактотропін, його ще називають лютеотропіном, лактогенним чи мамотропним гормоном) виділяється гіпофізом лише після утворення жовтого тіла. Він володіє багатосторонньою дією – стимулює ріст і секреторну діяльність жовтого тіла, ріст молочних залоз і лактацію, діючи безпосе-редньо на епітелій альвеол молочної залози, попередньо підготований естрогенами чи прогестероном. Пролактин є синергістом лютропіну і додатково стимулює розвиток жовтих тіл та утворення прогестерону. Інкрецію пролактину стимулюють естрогени, а прогестерон – її гальмує.
Під час вагітності секреція пролактину утримується на одному рівні, після родів вона різко зростає. Це виділення його підтримується нервово-рефлекторним шляхом при ссанні, доїнні та масажі вим’я. При тривалому надходженні пролактину жовте тіло розростається і виділяє більше прогестерону, який пригнічує ріст фолікулів. Тому багато молочних корів (з добовим надоєм 25–30 кг) не приходять в охоту про-тягом перших трьох місяців лактації внаслідок створення у головному мозку лакта-ційної домінанти.
Дві гонадотропні функції гіпофіза – продукція фолітропіну і лютропіну з одного боку і пролактину – з іншого, контролюються різним чином. Перша перебуває під гіпоталамічним контролем, друга є саморегулюючою функцією передньої долі гіпо-фізу, що проявляється при припиненні дії на нього гіпоталамуса.
Фолітропін діє на статеву систему лише через яєчник. Введення фолітропіну ка-строваним самкам не викликає морфологічних змін у їх геніталіях.
У самців гонадотропіни виділяються безперервно, а у самок – у певному ритмі, циклічно.
Характерною рисою статевої системи є періодичність оваріальної функції (до-зрівання фолікулів, секреції естрогенів, овуляції, існування жовтого тіла), вагітності, родів, лактації.
Зовнішніми подразниками, що стимулюють статеву функцію тварин, є світловий та температурний режим, рівень годівлі, а внутрішніми – концентрація статевих гор-монів. При відсутності цих модулюючих впливів відбувається тонічне (ациклічне) виділення високих концентрацій фолітропіну і лютропіну. Знизити цей рівень і пере-ключити секрецію гонадотропінів на циклічний тип можуть лише стероїдні гормони яєчника та інгібін.
Фізіологічні особливості статевих органів свійських тварин
Фолітропін та лютропін, діючи на фолікул, перш за все викликають внутрішньо-клітинні ферментні реакції. Таким ферментом у клітинній мембрані є аденілциклаза. Вона служить посередником між гонадотропним гормоном та внутрішньоклітинними ферментативними системами.
Секреція гонадотропінів гіпофізом регулюється за принципом зворотного зв’яз-ку з концентрацією у крові статевих гормонів. Встановлюється динамічна рівновага між секрецією гіпоталамічних нейрогормонів (Гн–РГ), гіпофізарних гонадотропінів та секрецією статевих гормонів. Статеві гормони, залежно від їх концентрації та спів-відношення з іншими гормонами, пригнічують або стимулюють продукцію відповід-ного тропного гормону. М. М. Завадовський назвав це плюс – мінус взаємодією, що пізніше отримало назву механізму зворотного зв’язку. Головним місцем дії стероїдів у механізмі зворотного зв’язку є гіпоталамус.
Гормони, що беруть участь у регуляції статевої функції, відзначаються чіткою специфічністю і дистанційністю дії.
Гонади (яєчники та сім’яники) є третьою ланкою у регуляції статевої функції, яку вони здійснюють через статеві гормони. Хімічні дослідження останніх років по-казали, що, не дивлячись на різниці у біологічній дії, хімічна структура чоловічих і жіночих статевих гормонів досить близька: це чотирикільцеві стероїдні сполуки типу фенантрену, одні з яких (естрадіол, естрон і естріол) мають фенолову чи ОН-групу, а другі (прогестерон і тестостерон) не мають цієї групи. У організмі тварин статеві гормони зв’язані з органічними кислотами.
| β |
Естрогени, андрогени, прогестерон та релаксин синтезуються у тварин обох ста-тей, лише у різних кількостях.
Усі стероїди синтезуються з холестеролу (що утворюється клітинами внутрішньої оболонки фолікула з ацетату, або ж заносяться сюди плазмою крові) через прегнено-лон і мають спільну основну структуру. Під впливом наявної тут дегідрогенази пре-гненолон окислюється до прогестерону, а тоді перетворюється у андрогени (дегідро-епіандростерон, андростендіон та тестостерон). Вважають, що андрогени відіграють певну роль у самок в підтриманні лібідо, крім того, вони попереджують атрезію фо-лікулів. Згодом, через серію ферментативних реакцій (ароматизація), наявні у стінці фолікула андрогени перетворюються в естрогени.
Перетворення наявних у стінці фолікула андрогенів у естрогени відбувається під впливом фолітропіну.
При розладах цього процесу в організмі самки замість естрогенів може зростати концентрація андрогенів з відповідними змінами в організмі. Крім того, невеликі кіль-
Розділ 2
кості естрогенів утворюються у жовтому тілі та плодових оболонках у другій полови-ні вагітності. Гранульозні клітини преовуляторного фолікула секретують також про-стагландини Е2та F2α, які відіграють певну роль у розриві фолікула, а також лютеолізі жовтого тіла у приматів. У жовтому тілі синтезується також релаксин та окситоцин.
Слід мати на увазі, що гонадотропіни різних видів ссавців відрізняються один від одного за хімічним складом та антигенними властивостями, хоча за біологічною дією тут відмінностей немає.
Вміст естрадіолу в сироватці крові змінюється протягом циклу. Так, у телиць він починає збільшуватися за три дні до початку тічки, досягає максимуму за 12 годин до неї; у крові овець концентрація естрадіолу також підвищується за три дні до початку тічки, досягаючи максимуму в кінці тічки. На 4-й та 9-й день циклу спостерігаються додаткові піки концентрації гормону. Подібні зміни мають місце і у крові свиней. У них відмічено також підвищення концентрації гормону на 26–30-й день поросності та перед опоросом.
Рівень тестостерону у самок у 10–15 разів нижчий, ніж у самців. Прогестерон продукується також інтерстиціальними клітинами сім’яників, а естрогени – клітина-ми Сертолі, проте фемінізації не наступає, оскільки у них немає рецепторів для естро-генів; вони швидко інактивуються та піддаються ферментативному розкладу.
Накопичуючись у фолікулярній рідині такрові, естрогени активують нервову сис-тему самки і викликають формування у ній статевої домінанти. У самки виникає ста-теве збудження, що переходить згодом у охоту. У статевих органах естрогени діють лише на провідні шляхи – (матку і піхву), викликаючи їх гіперемію, секрецію і про-ліферацію, розслаблення шийки матки і звільнення її від слизу, підвищуючи їх чутли-вість до окситоцину і підготовляючи геніталії самки до подальшої дії прогестерону. Крім того, естрогени посилюють розвиток системи проток молочної залози, особливо при статевому дозріванні молодих тварин, проте вони гальмують процес лактації.
Біологічна дія естрогенів багатогранна. Під впливом фолітропіну стимулюється ріст фолікулів, який носить неоднорідний характер. Серед них виділяється домінант-ний, що продукує наростаючі кількості інгібіну, фолікулярного регуляторного проте-їну (ФРП) та естрадіолу. Інгібін та естрадіол, діючи на гіпофіз, сприяють зниженню секреції фолітропіну і блокують цим появу нових великих фолікулів, а ФРП пригнічує синтез естрадіолу другорядними фолікулами і у них наступає атрезія. Проникаючи у кров’яне русло, естрогени адсорбуються білками, доносяться до органів та тканин, звільняються від білка, проявляють свою дію залежно від концентрації: малі дози сти-мулюють ріст фолікулів, поділ клітин, тоді як великі дози пригнічують виділення фо-літропіну у передній долі гіпофіза і викликають атрофічні зміни в тканинах яєчника.
Анаболічна дія естрогенів яскраво проявляється при гіперестрогенізації організму при кістозному переродженні яєчників, коли у тварини спостерігають тривалу німфо-манію та зміни зовнішнього вигляду самки. Впливаючи на ліпідний обмін, активуючи імунні реакції та фагоцитоз, естрогени підвищують захисні сили організму.
Фізіологічні особливості статевих органів свійських тварин
Заносячись з кров’ю у печінку, естрадіол перетворюється у менш активні естрон і естріол, які нагромаджуються у крові та сечі самок і виділяються з калом; концентра-ція їх починає наростати з 45-го дня, а у кобил і корів – з 2–3-го місяця вагітності. Про-те охоти вони не викликають, оскільки високі концентрації прогестерону пригнічують виділення лютропіну і дозрівання фолікулів не завершується. Крім того, ці естрогени не завжди бувають у активній формі, а зв’язуються з глюкуроновою кислотою.
Під час вагітності естрогени обумовлюють ріст матки та морфологічну і функці-ональну перебудову її нервово-м’язового апарату – у м’язах матки тут синтезуються скоротливий білок (актоміозин), макроергічні фосфати (АТФ), вуглеводи (глікоген), електроліти (кальцій, калій) й інші біологічно активні речовини; зростає кровообіг і підвищується парціальний тиск кисню, поліпшуються окисно-відновні процеси у міометрії і т. д., тобто, вони готують нервово-м’язовий апарат матки до активної ско-ротливої діяльності. Під кінець вагітності вміст естрогенів уже переважає над прогес-тероном, що робить матку чутливою до окситоцину і забезпечує ритмічні скорочення м’язів під час родів.
Регуляція гормональної функції гонад здійснюється ступенево за кірково-гіпота-ламо-гіпофізарним типом, а також безпосередньо за типом кора – гонади, гіпотала-мус – гонади.
У 40-х роках минулого століття встановлено, що естрогенними властивостями во-лодіють і синтетичні сполуки, які не мають нічого спільного з яєчниками, фолікуляр-ними та плацентарними гормонами. Багато з них за своєю активністю значно пере-вищують фолікулярні гормони, чому вони отримали широке практичне застосування (синестрол, гексаестрол, вітестрол, дигідростільбестрол).
Гормонижовтоготіла. Жовте тіло функціонує як тимчасова залоза внутрішньої секреції, виробляючи гормон прогестерон (прогестин, лютеїн, лютеостерон). Завданням прогестерону є “закінчити те, що розпочали естрогени”. Крім жовтого тіла прогестерон виробляється також, правда, у значно меншій кількості, у клітинах гранульози яєчника, корі наднирників та плаценті, сім’яниках самців. У печінці прогестерон інактивується і перетворюється у прегнандіол, прегнанол, прегненолон, глюкуроновий ефір.
Вплив естрогенів і прогестерону на статеві органи самки має як синергічний, так і антагоністичний характер. З одного боку, прогестерон діє на слизову оболонку матки лише після підготовчого впливу естрогенів, з іншого, він пригнічує скорочення матки, тоді як естрогени їх стимулюють.
Дія прогестерону проявляється у трьох напрямках: а) підготовці ендометрію до імплантації ембріона і збереженні плода протягом всієї вагітності (при нестачі гор-мону вагітність переривається); б) гальмуванні росту міометрію, стимулюванні його гіпертрофії і гіперплазії; в) стимуляції росту альвеол молочної залози.
Під впливом прогестерону слизова оболонка матки трансформується, у ній за-кінчується проліферативна фаза і розвивається секреторна, що супроводжується роз-рихленням епітелію ендометрію, збільшенням вмісту глікогену у його цитоплазмі та секреті маткових залоз.
Розділ 2
Пригнічуючи виділення передньою часткою гіпофіза лютропіну, прогестерон гальмує дозрівання вторинних фолікулів у яєчниках – овуляцію, а у молочних залозах він викликає проліферацію епітелію у їх аценозній частині.
Концентрація прогестерону у крові починає зростати з 4-го дня циклу, досягаючи максимуму на 7–8 день, а за деякими авторами на 10–12-й день, з 14–15-го дня вона знижується і особливо різко перед наступною охотою.
Імпульс для секреції прогестерону (як і естрогенів) надходить з кори головно-го мозку. Термін функціонування жовтих тіл контролюється маткою, у епітеліальних клітинах якої синтезується біологічно активна речовина лютеолітичної дії – проста-гландин F2α, який по кровоносних судинах доноситься до яєчника, акумулюється у лютеальній тканині і викликає швидку її регресію.
Гормон жовтого тіла і його синтетичні замінники останнім часом широко застосо-вуються для вкорочення сильно вираженої тічки, попередження спонтанних абортів, синхронізації охоти і т. п.
Чоловічістатевігормони(андрогени). Сюди належать андростерон, дигідро-андростерон, тестостерон і споріднені з ними речовини. Вважають, що первинними статевими гормонами є тестостерон, а андростерон і дигідроандростерон – продукти біологічних змін його в організмі. У хімічному відношенні андрогени належать до тієї ж групи стероїдних гормонів, що й естрогени. Вони викликають у молодих самців розвиток вторинних статевих ознак і покручених каналиків сім’яника, а у статеводоз-рілих тварин – статевий потяг і впливають на вираженість вторинних статевих рис. При розладах регуляції статевої функції у самок андрогени можуть викликати в них маскулінізацію.
Утворюються андрогени у сім’яниках та корі наднирників, піддаються в організмі метаболічним перетворенням і виводяться з сечею у вигляді 17-кетостероїдів.
Тестостерон відіграє роль не лише у репродуктивній функції. Він стимулює також синтез білка, чому самці мають більшу масу тіла, ніж самки. Крім того, тестостерон може підсилювати еритропоез.
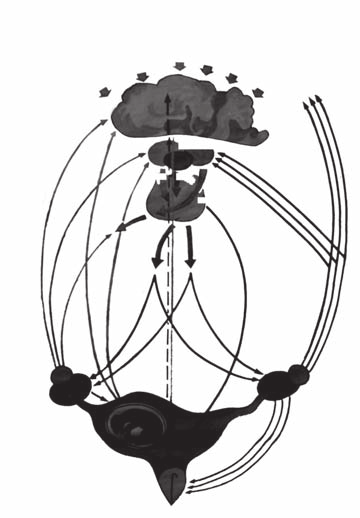
Схему регуляції статевої функції у самок можна уявити собі так: імпульси, що над-ходять у кору головного мозку від зовнішнього середовища (через органи слуху, зору, нюху, теплові таінші подразники), особливосильні у присутності самця, передаються по центробіжних нервах у відповідні центри кори головного мозку, сумуються тут з імпульсами від внутрішніх органів, переробляються і передаються у гіпоталамус, викликаючи в ньому стан підвищеної або ж зниженої функціональної активності. У першому випадку в супраоптичних та паравентрикулярних ядрах гіпоталамуса виро-бляється нейросекрет (рилізінг-фактор), який, поступаючи у гіпофіз, стимулює виді-лення ним фолітропіну. Останній поступає у кров і, доносячись до яєчника, викликає ріст і дозрівання фолікулів, що супроводжується синтезом у фолікулярних клітинах естрогенів. Діючи на хеморецептори та аналізатори головного мозку, вони виклика-ють у тварини тічку, охоту та загальне збудження (рис. 18).
Фізіологічні особливості статевих органів свійських тварин
 І
І
гіпоталамус
гн рф СТ. ОХОТА
гіпофіз
ЕСТРОГЕНИ
 ЕСТРОГЕНИ
ЕСТРОГЕНИ
жовте тіло фолікул
тічка Рис. 18. Схема регуляції статевого циклу.
Розділ 2
Високі концентрації естрогенів у крові пригнічують виділення фолітропіну і сти-мулюють виділення лютропіну, який викликає овуляцію і лютеїнізацію зернистої обо-лонки фолікула. Жовте тіло, що виникло після овуляції, виділяє прогестерон, який гальмує дальше виділення лютропіну, стимулює лютеотропну функцію і не перешко-джає секреції фолітропіну, а у вагітних самок стимулює лютеотропну функцію гіпо-фіза. На цьому фоні зростає секреція прогестерону.
При відсутності запліднення ендометрій виробляє в кінці статевого циклу люте-олітичний фактор (ПГ-F2α), що викликає регресію жовтого тіла і статевий цикл по-новлюється.
У великої рогатої худоби перша після овуляції хвиля росту фолікулів завершуєть-ся їх атрезією, що стимулює нову хвилю росту фолікулів. Протягом статевого циклу у них буває дві, а то й три хвилі росту фолікулів. Внаслідок останньої хвилі виникає домінантний фолікул, який і овулює.
Для дозрівання фолікула, настання тічки та овуляції необхідне співвідношення фолітропіну до лютропіну від 4: 1 до 100: 1.
Гормони яєчників, з одного боку, проявляють вплив на периферійні статеві орга-ни, підготовляючи їх до імплантації ембріона, з другого боку, вони беруть участь у регуляції функції системи гіпоталамус – гіпофіз за принципом зворотного зв’язку.
Підсумовуючи сказане, циклічний характер статевої функції самок можна пояснити:
Ø наявністю у центральній нервовій системі своєрідного біологічного годинни-ка чи як ще його називають циклічного центру у вигляді гіпоталамусу;
Ø періодичним виділенням цим центром рилізінг – гормонів;
Ø відповідною зміною секреції гіпофізом фолітропіну та лютропіну;
Ø послідовною зміною в яєчнику секреції естрогенів та прогестерону (фолікулі нової та лютеїнової фаз статевого циклу);
Ø наявністю системи зворотного зв’язку у ланцюгу гіпоталамус – гіпофіз – яєч-ники та формуванням у корі головного мозку статевої домінанти, як ознаки готовності організму до запліднення.
Якщо статевий цикл завершується заплідненням, то у корі головного мозку фор-мується материнська домінанта, яка в кінці вагітності проявляється як родова, а зго-дом – лактаційна домінанта.
Крім згаданих гормональних впливів, на функцію статевої системи впливають ті-реотропний, адренокортикотропний, соматотропний, а також негіпофізарні гормони епіфіза, наднирників, щитовидної та інших залоз.
Як відомо, під час вагітності роль ендокринної залози з багатогранною дією ви-конує плацента. У її ворсинках виробляються значні кількості естрогенів та прогесте-рону, а також гонадотропний гормон – сироватковий чи плацентарний (у жінок хорі-альний) гонадотропін.
У кобил сироватковий гонадотропін з’являється у сироватці крові і лімфі з 4-го дня жеребності, коли матка готується до імплантації зародка. Максимальної концен-
Фізіологічні особливості статевих органів свійських тварин
трації (80–160 МО/л) гонадотропіни досягають на 50–100-й день жеребності, опісля концентрація їх швидко падає. Укобил сироватковий гонадотропін не проходить крізь нирковий фільтр і тому затримується у крові, тоді як у жінок він проникає у сечу і на-копичується там у великій кількості. Під час вагітності плацентарний гонадотропін спочатку доповнює дію гіпофізарних гонадотропінів, а опісля заміняє їх.
Важливе значення для відтворної функції має також гормон задньої долі гіпофіза окситоцин. Він стимулює скоротливу функцію матки і є однією з ланок механізму родової діяльності. Істотна роль йому належить в регуляції лактації.
Ще у 60-х роках ХХ ст. було відмічено взаємозв’язок між станом матки та три-валістю функціонування жовтого тіла. На цій підставі було зроблено висновок, що у рогах матки виробляються лютеолітичні речовини місцевої дії. Дослідження останніх років показали, що цю роль виконують простагландини (ПГ).
Вперше простагландини були відкриті в спермі. У 1930 р. американські гінекологи Курцрок і Льоб повідомили, що матка жінки реагує на введення сперми скороченням чи розслабленням. У 1935 р. шведський учений Ейлер довів, що тонізуючим впли-вом на гладку мускулатуру володіє ліпідний екстракт із сперми і назвав його проста-гландином (вважаючи, що він утворюється в простаті), а у 1957 р. простагландин F 2α було екстраговано за допомогою фосфатного буфера з тканинипередміхурової залози барана. При структурному аналізі простагландину в його білкових ланцюгах було ви-явлено подвійний зв’язок та α-конфігурацію 9-ї гідроксильної групи. Ці особливості було відображено в назві сполуки. За це відкриття С. Бергстрем був удостоєний Но-белівської премії.
Нині відомо, що простагландини синтезуються у клітинних мембранах більшості тканин ссавців.
Простагландин F2α володіє вираженою лютеолітичною дією, вкорочуючи наполо-вину функцію жовтого тіла, що супроводжується зниженням у крові рівня прогесте-рону та підвищенням концентрації стероїдів. Він також викликає початок родів та швидке скорочення судин пупкового канатика.
Дослідження останніх років значно розширили розуміння суті статевого циклу та його регуляції. Згідно П. К. Анохіна усі функціональні системи є саморегульованими, а за К. В. Судаковим, зміна поведінки тварин визначається внутрішніми потребами їх організму. За теорією функціональних систем основу статевого циклу складають періодично повторювані циклічні зміни у системі гіпоталамус – гіпофіз – яєчники в зв’язку з ростом та дозріванням фолікулів, овуляцією, формуванням і функціонуван-ням жовтого тіла. Сюди також входять циклічні зміни матки, інших систем організму та поведінки тварини.
Запорукою нормального прояву статевого циклу є синхронність дії основних ре-гуляторних систем, що контролюють процес розмноження (А. Г. Нєжданов).
Гонадотропні гормони гіпофізу – фолітропін, що контролює ріст фолікулів та синтез естрогенних гормонів, а також лютропін, що забезпечує дозрівання фолікулів,
Розділ 2
овуляцію, синтез прогестерону та естрогенів, проявляють свою активність лише при певному їх співвідношенні.
З дозріванням у яєчниках фолікулів зростає виділення ними естрогенних гормо-нів, які, в свою чергу, у передовуляційну фазу активують виділення гіпоталамусом Гн–РГ, підвищують чутливість до нього аденогіпофізу, у якому різко зростає секреція ЛГ, що забезпечує дозрівання і овуляцію фолікула.
З утворенням жовтого тіла і секрецією ним прогестерону за принципом зворот-ного зв’язку гальмується дальше виділення ЛГ. Під впливом малої частоти виділення Гн–РГ та ФСГ стимулюється ріст фолікулів. Поява домінантного фолікула супрово-джується інтенсивною секрецією естрадіолу та інгібіну. Останній гальмує ріст інших фолікулів. Така черговість проявляється декілька раз протягом статевого циклу, що кожен раз завершується атрезією домінуючого фолікула, крім останнього, передову-ляційного.
Стимулом частоти пульсації Гн–РГ, а за ним виділення ЛГ є зниження вмісту прогестерону, а високі концентрації естрадіолу, навпаки, стимулюють виділення ЛГ (Imakawa з співр.).
В регуляції статевої функції гіпофізу беруть також участь такі нейромедіатори гіпоталамуса, як норадреналін, дофамін та серотонін, а також продуковані у ЦНС опіоїдні пептиди енкефаліни, ендорфіни та динорфіни, зокрема у передачі нервових імпульсів.
Формування феноменів стадії збудження статевого циклу відбувається під впли-вом статевих гормонів лише при певному естрогено-прогестероновому та андрогено-прогестероновому співвідношеннях (А. Г. Нєжданов). Перша хвиля росту фолікулів у яєчниках у лютеїнову фазу статевого циклу не викликає змін у поведінці тварин. Лише друга – третя хвиля росту фолікулів, на фоні регресії жовтого тіла, зниження концентрації прогестерону, підвищення секреції андрогенів і естрогенів приводить до формування стадії збудження статевого циклу та її феноменів. При цьому пік концен-трації естрогенів припадає на період яскраво вираженої тічки та статевого збудження, а тестостерону – на період прояву охоти.
Поряд з цим значну роль у регуляції статевої функції відводять тиреоїдним гормо-нам, гормону епіфіза мелатоніну, пролактину і іншим біологічно-активним речовинам. Штучна регуляція статевого циклу. Знання особливостей статевого циклу на-штовхнуло вчених на ідею штучної регуляції відтворювальної функції, практичне за-стосування таких заходів, як стимуляція багатоплідності, синхронізація охоти, віднов-лення статевої циклічності, корекція часу овуляції, часу настання статевого сезону. У 1934 р. довів Лопирін А. І. можливість стимуляції суперовуляції, що опісля підтверди-ли М. М. Завадовський, А. Л. Падучева, Роусон, Гердон, Еверілл, Робінсон та ін. Ши-рокого практичного застосування при цьому набули сироватка крові жеребних кобил (СЖК) та очищений її препарат (ГСЖК) – сироватковий гонадотропін, а також виго-товлений з сечі жінок – хоріальний гонадотропін (ХГЛ). Вказані гонадотропіни володі-ють фізіологічною дією ФСГ і ЛГ, причім в отриманій з 50-го по 70-й день жеребності
Фізіологічні особливості статевих органів свійських тварин
сироватці жеребних кобил звичайно переважає ФСГ, а пізніше – ЛГ. Хоча сироватко-вий гонадотропін володіє більше дією ФСГ, а хоріальний – ЛГ. Отримана на 50–100-й день вагітності СЖК використовується для стимуляції дозрівання фолікулів.
Застосування у 1939 р. СЖК у Середній Азії на 22 тисячах вівцематок дозволило підвищити вихід ягнят до 150–200 від кожної сотні вівцематок і отримати додатково 7 тис. ягнят. З тих пір цей метод стали широко використовувати у тваринництві, осо-бливо для отримання каракульських двійнят, де більше половини приплоду забива-ється на смушки. На жаль, масові обробки тварин без врахування їх фізіологічного стану, особливо на фоні неповноцінної годівлі привели з часом до погіршення якості смушків, зниження їх товарної цінності на міжнародному ринку і врешті ці обробки овець припинили.
У Великобританії дещо видозмінили цю методику – спочатку, протягом 8-ми діб, овець обробляють прогестероном (вкладаючи у їх піхву просочені прогестероном пе-сарії), а тоді застосовують СЖК, а в Австралії застосування СЖК поєднують з вну-трішньом’зовою ін’єкцією на 12–13-ту добу статевого циклу люліберину. Є й інші модифікації.
Корови менш схильні до багатоплідності. Найефективнішим методом стимулю-вання у них багатоплідності виявилася 4-разова ін’єкція з 12-годинним інтервалом фолітропіну у дозі 20 мг на 10–12-ту добу статевого циклу. Проте, у 34 % оброблених корів наступали аборти на 7–10-ту добу тільності (головно, при вагітності 3–4 плода-ми); у 4–5 разів зросли випадки мертвонароджень, біля 92 % теличок народжувалися фрімартинами; удвічі зросла смертність новонароджених телят; частими були випад-ки важких родів та затримання посліду (М. І. Полянцев).
Стимулювання багатоплідності у свиней також не дало бажаних наслідків. Чи-сельність приплоду зростала всього на 1–1,5 поросяти.
Удосконалення методів біотехнологічного керування відтворенням дозволило з часом успішно застосовувати й інші гормональні препарати. Особливо ефективними тут виявилися гіпофізарні гонадотропіни – ФСГ та його препарати, які дозволяють довести рівень овуляції у одноплідних тварин до 5–25 і більше.
Головною методичною помилкою більшості робіт із стимуляції статевої функції є масове застосування препаратів на великому поголів’ї тварин без врахування у них фаз статевого циклу. Статеві центри і генеративні органи тварин, як вже зазначалося, по-різному реагують на гонадотропні та гонадальні гормони у різні фази циклу. Як встановив А. І. Лопирін, у передовуляційну фазу яєчники чітко реагують на виділю-ваний в кров фолітропін і лютропін, а тканини родостатевих шляхів відповідають тіч-ковими змінами на продуковані у яєчниках естрогени. В післяовуляційну фазу яєчни-ки втрачають чутливість до фолітропіну і реагують лише на лютропін чи пролактин, а матка стає толерантною до естрогенів, зате вона робиться чутливою до прогестерону. Нарешті, в лютеїнову фазу ні гонадотропні, ні естрогенні гормони не викликають по-мітних морфологічних змін у яєчниках і статевих шляхах, а матка продовжує функці-онувати під впливом прогестерону.
Розділ 2
Тому застосування гонадотропних і естрогенних препаратів у передтічкову ста-дію буде підсилювати дію ендогенних гормонів, тоді як під час охоти вони лише сти-мулюватимуть появу гіпертрофічних фолікулів, а то й лютеїнізацію їх без овуляції. Застосування гонадотропінів у лютеїнову фазу приводить до затримання інволюції циклічних жовтих тіл.
Нервові статеві центри реагують на застосовані обробки в ширшому діапазоні. Тому введенням естрогенних препаратів можна викликати ановуляторний стате-вий цикл.
Слід також пам’ятати, що у яєчниках завжди є певна кількість фолікулів різного ступеня зрілості, внаслідок чого суперовуляція часто супроводжується виділенням значної кількості яйцеклітин різної біологічної цінності.
Нарешті, застосування СЖК у лютеїнову фазу циклу не рідко приводить до пер-систенції і кістозного переродження жовтих тіл, що також несприятливо позначається на відтворювальній функції самок.
Широкого застосування у тваринництві набув метод синхронізації статевої ци-клічності: у скотарстві при трансплантації ембріонів, у м’ясному скотарстві та ві-вчарстві – для одночасного сезонного осіменіння тварин; на свинокомплексах – для організації турових опоросів.
З цією метою використовують різні препарати прогестагенів, які або ін’єктують тваринам протягом певного часу (6–15 днів), або вводять їх у вигляді піхвових там-понів, спіралей та ін., або згодовують з кормом. Під впливом наявного у цих препара-тах прогестерону гальмується дозрівання антральних фолікулів, а через декілька днів після припинення обробки у тварин виникають ознаки статевого збудження. Широко використовуються у молочному скотарстві для синхронізації статевої функції пре-парати простагландинів – F2α та їх синтетичні аналоги. Дворазові ін’єкції цих препа-ратів з інтервалом 10–11 днів забезпечують синхронну появу ознак статевого циклу у більшості тварин.
Значну увагу у 80-х роках минулого сторіччя приділяли синхронізації охоти у тва-рин. При цьому відпадала необхідність щоденного контролю за охотою та овуляцією; це дозволяло планувати час осіменіння тварин та отримання від них приплоду, поліп-шувалася організація осіменіння.
У практиці тваринництва сформувалися два напрямки синхронізації охоти: 1) пригнічення на певний час росту оваріальних фолікулів у групи тварин з поновлен-ням фолікулогенезу після припинення дії препарату; 2) одночасне викликання регре-сії жовтих тіл у групи тварин.
У першому випадку тваринам вводять прогестерон протягом часу, характерного для фази секреторної активності жовтого тіла, що обумовлює інволюцію жовтих тіл. Метод базується на здатності прогестерону активізувати через механізм зворотного зв’язку біосинтез фолітропіну. При цьому також зростає чутливість до фолітропіну клітин-мішеней у тканинах яєчників. Після припинення введення препарату у яєчни-
Фізіологічні особливості статевих органів свійських тварин
ках усіх тварин розпочинається одночасний ріст передовуляційних фолікулів, синх-ронно наступає охота та овуляція.
Прогестерон можна задавати тваринам впродовж 12 днів з кормом, у вигляді вагі-нальних песаріїв, силіконових спіралей, вводити підшкірно. Через 48–72 години піс-ля припинення обробки тварин осіменяють.
Проте запліднюваність корів у синхронізовану охоту низька.
У вівчарстві для синхронізації охоти застосовують задавання per os протягом 14 днів прогестагенного препарату ацетату мегестролу у суміші з кормовою сіллю (1: 5000). Через добу після припинення задавання препарату вводять тваринам по 1000 І.О. СЖК і ще через добу осіменяють.
Для синхронізації охоти у свинарстві широко використовували задавання з кор-мом протягом 20 днів по 100 мг нестероїдного препарату металібур, що блокує синтез та виділення фолітропіну. Через добу після останнього задавання препарату вводили тваринам СЖК, ще через 4 доби – ХГЛ і через 24 години осіменяли. Запліднювалися до 90 % свиноматок, проте у випадку задавання препарату поросним свиноматкам серед народжених поросят було багато виродків, тому застосування цього препарату припинили.
У скотарстві для синхронізації охоти найчастіше застосовують препарати проста-гландину F2α, які ін’єктують коровам, незалежно від стану геніталій, з 40–45-го дня після отелення двічі, з інтервалом 11 днів. Через 80 та 92 години після повторного введення препарату тварин осіменяють.
Ефективним засобом корекції статевої функції є синтетичний аналог рилізінг-гор-мону – сурфагон, що випускається в ампулах і флаконах. Мінімальна фармакологічна доза для корів, що викликає виділення гіпофізом фолітропіну та лютропіну – 100 мкг, терапевтична – 10–50 мкг. Вже у перші години після ін’єкції препарату стимулюєть-ся виділення гіпофізом гонадотропних гормонів. Препарат вводять на 14–20-й день після отелення у дозі 10 мл (50 мкг). Це обумовлює поновлення у тварин статевої циклічності, але їх осіменяють лише у другу статеву охоту.
Другим аналогом Гн–РГ можна назвати диригестран, що застосовується у дозі 100–250 мкг (2,5–6,25 мл) для стимулювання статевої циклічності у високопродук-тивних корів після отелу, при гіпофункції та кістах яєчників.
З такою ж метою застосовують фертирелін (100 мкг), гонадорелін (0,5 мг), бусе-рилін (10–20 мкг).
Сурфагон використовують також для стимулювання статевої функції у свинома-ток, ін’єктуючи їм на другий день після опоросу1500–2000 І. О. СЖК і через 56 годин 2 мл (10 мкг) сурфагону.
Сурфагон можна також застосовувати для підвищення заплідненості при штуч-ному осіменінні. З цією метою препарат вводять коровам у дозі 2 мл, внутрішньо-мускульно, на початку тічки, свиноматкам – 3 мл. Овуляція наступає у корів через 40 годин, у свиней – через 39–46 годин після ін’єкції препарату. Заплідненість самок зростає на 10–20 % (М. І. Полянцев).
Розділ 2
2.2. Статева та фізіологічна зрілість самців, сперматогенез
Статева та фізіологічна зрілість у самців наступає пізніше, ніж у самок.
Статеве дозрівання самців включає ріст статевих органів, появу статевих рефлек-сів та початок сперматогенезу. Першими ознаками процесу статевого дозрівання є поява у самців статевого потягу та парувального рефлексу, які з’являються, залежно від породи, у бугайців у 5–6-місячному віці та рефлексу ерекції, що з’являється у 7–9 місяців.
Початок активного сперміогенезу, з появою у придатку сім’яника дозрілих спермі-їв, припадає на 7–8,5 місяців. Перша еякуляція у них проявляється у 8,5–11 місяців.
Протягом часу від першого статевого потягу до першої еякуляції відбувається ріст та розвиток прутня, його голівка звільнюється від препуція і набуває остаточної форми. Сім’яники у нього вже функціонують як орган з подвійною генеративною та ендокринною функцією.
Розвиток статевого інстинкту у самців супроводжується відповідними змінами їх екстер’єру.
Важливо запам’ятати і таке: від статевого дозрівання і до племінного викорис-тання плідника проходить певний відрізок часу, тим довший, чим пізніше починають використовувати самця. Доведено, що на час початку статевого використання бугаїв у віці 16–18 місяців, кнурів – у віці 11–12 місяців у частини з них уже бувають різні статеві збочення, зокрема, онанізм, який викликає статеве виснаження плідників, що зберігається на все життя. У інших плідників, особливо у кнурів та баранів, розви-вається імпотенція. При пізньому початку статевого використання у плідників важко виробляються рефлекси на штучну вагіну. Отже, як з фізіологічної, так і з господар-ської точки зору важливо правильно визначити час початку статевого використання плідника.
Таблиця 5 Строки статевого та фізіологічного дозрівання самців окремих видів тварин
| Вид тварин | Статева зрілість | Фізіологічна зрілість |
| Бугай | 6–9 міс. | 16–18 міс. |
| Баран | 6–8 міс. | 15–18 міс. |
| Цап | 7–8 міс. | 1,5–2 роки |
| Кнур | 5–6 міс. | 10–11 міс. |
| Жеребець | 12–15 міс. | 3–4 роки |
| Верблюд | 2,5–3 роки | 4–5 років |
| Пес | 6–8 міс. | 12–14 міс. |
| Кріль | 4–5 міс. | 4–14 міс. |
| Кіт | 4–5 міс. | 4–14 міс. |
Фізіологічні особливості статевих органів свійських тварин
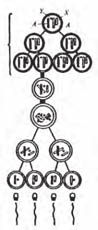
Сперматогенез (або спермогенез) – процес утворення сперми, що включає спер-міогенез – процес утворення та дозрівання чоловічих статевих клітин – сперміїв*, який розпочинається у покручених каналиках сім’яника і завершується у придатку сім’яника та плазмогенез – утворення плазми сперми.
Сперміогенез у сільськогосподарських тварин ділять умовно на 4 стадії: розмно-ження (поділу), росту, дозрівання і формування. С. У. Солсбері і Н. Л. Ван Демарк об’єднують перші три стадії в одну фазу – сперматоцигогенез, а сперміогенезом вони називають лише останню стадію.
Якщо розглянути під мікроскопом гістопрепарат сім’яника, то можна побачити, що на базальній мембрані покручених каналиків сім’яників розташовані два види клі-тин. Одні з них – великі з великим ядром і протоплазмою, що виступає у просвіт кана-лика у вигляді полум’я свічки. Це – соматичні клітини Сертолі, які виконують опірну і трофічну функцію. Вважають, що високий вміст глікогену в цитоплазмі синцитія цих клітин служить джерелом енергії для сперміїв на останній стадії їх формуван-ня. Між клітинами Сертолі розташовані у декілька шарів сперміоутворюючі клітини, що перебувають на різних стадіях сперміогенезу. Розміщений на базальній мембрані каналика нижній шар порівняно менших клітин представлений сперміотворними клі-тинами – сперматогоніями. Вони містять відносно велике ядро і характерну для сома-тичних клітин кожного виду кількість хромосом (наприклад: у бугая – 60, цапа – 60, жеребця – 66, барана – 54, кнура – 40).
Перші ознаки сперміогенезу самців можна спостерігати ще в ембріональний пе-ріод, з часу статевої диференціації, коли первинні статеві клітини у складі зародків гонад дають початок сперматогоніям. Проте інтенсивне утворення сперміїв розпочи-нається з настанням статевої зрілості самців (у бугая – в 6–9 місяців). У цей час спер-матогонії починають розмножуватись і розташовуються вже у каналику в 3–7 шарів.
Стадія розмноження сперматогоній відбувається мітотичним шляхом, причому одна сперматогонія може дати до 10 і більше поколінь дочірних клітин, рівноцінних материнській за об’ємом, морфологією та хромосомним набором. Коефіцієнт розмно-ження сперматогоній у бугая та барана дорівнює від 8 до 16, у кнура – до 24.
Стадія росту. Одна з новоутворених клітин залишається на місці (сперматого-нія А), а друга (Б) просувається у напрямку просвіту каналика, вступає в стадію рос-ту – збільшується в об’ємі і перетворюється у сперматоцит 1-го порядку, що переви-щує сперматогонію за розміром приблизно у чотири рази. У ядрі сперматоцита від-бувається інтенсивний синтез ДНК, кон’югація хромосом.
Стадія дозрівання характеризується перш за все внутрішніми змінами хромо-сомного матеріалу і двома послідовними поділами сперматоцитів – редукційним та
* В різний час чоловічі статеві клітини називали по-різному: Левенгук, побачивши вперше під мікроскопом ці рух-ливі клітини, назвав їх “анімалькулями” – звірятками, з часом з’явилися назви “сперматозоон” – від слів sperma (сім’я) та zoon (тварина) – сім’яна тварина; “сперматозоїд” – від тих же слів та idios (подібний) – подібний до сім’яної тварини; “спермій” – сім’яна клітина, “сім’яна нитка”, “живчик”. Найправильніше їх називати спермії, у медико-біологічній літературі широко використовується термін сперматозоїд. Терміни “сперматозоон”, “сім’яна нитка” та “живчик” не мають логічного підґрунтя.

 Розділ 2
Розділ 2
1 6
А
стадія розмноження
3 стадіяросту
Б
стадія дозрівання 6
10 стадіяформування
Рис. 19. Поперечний розріз сім’яного каналика (А) та схема сперміогенезу (Б):
1 – сперматогенний епітелій; 2 – сполучно-тканинна стінка канальця; 3 – просвіт канальця зі сперміями; 4 – зона сперматид; 5 – зона сперма-
тоцитів; 6 – зона сперматогоній; 7 – синтицій Сертолі.
екваційним, у результаті чого сперматоцит 1-го порядку ді-литься на два сперматоцити 2-го порядку, які в свою чергу діляться на чотири спермати-ди. Тобто з кожного сперма-тоцита І порядку утворюється чотири сперматиди.
Під час редукційного ме-йотичного поділу ядра хромо-соми об’єднуються попарно і, не розщеплюючись вздовж, розходяться до полюсів дочір-них клітин – сперматоцитів 2-го порядку, забезпечуючи їх половинним (гаплоїдним) набором хромосом. Еквацій-ний поділ (за типом мітозу) відбувається дуже скоро після редукційного, при цьому кож-на хромосома розщеплюєть-ся вздовж на дві половинки, здатні відновити втрачену по-ловину (редуплікація).
Сперматида, що при цьому утворилась, містить зернис-те ядро, у її цитоплазмі роз-міщена центросома, оточена оболонкою (центросферою чи ідісомою), навколо якої згру-пований у вигляді зерен чи пластинок апарат Гольджі; хон-дріосомний чи мітохондріаль-ний апарат виступає у вигляді дрібних паличок чи зерен.
Стадія формування від-бувається у верхівках прото-плазматичних відростків клі-тин Сертолі, що виступають у просвіт каналика, і характери-зується складними морфоло-гічними змінами сперматид.
80
Фізіологічні особливості статевих органів свійських тварин
Клітинне ядро сперматиди переходить у верхню її частину і вона приймає овальну форму. Розташовані біля переднього полюса ядра тільця Гольджі стають плоскими і утворюють над ним ковпачок. Між полюсом і ковпачком виникає вакуоля з розмі-щеною у ній ідіосомою (акросомною зерниною), яка переміщується до переднього полюса ядра і разом з рідиною вакуолі утворює акросому. Спорожніла ідіосома разом з апаратом Гольджі пересувається до протилежного полюса ядра і утворюють дві цен-тросоми – проксимальну (нижчу), що наближається до ядра, і дистальну (дальшу), розміщену на периферії сперматиди. Остання, давши початок вісьовий нитці джгутика спермія, відокремлює від себе центросомне кільце навколо цієї нитки і перетворюєть-ся з часом у основне кільце спермія, яке поступово пересувається разом з навколиш-ньою протоплазмою у напрямку хвостика. Ділянка вісьової нитки джгутика, розта-шована між центросомним і основним (базальним) кільцем, перетворюється згодом у тіло спермія. З мітохондріального матеріалу цитоплазми формується спіральна нитка, що обмотується навколо джгутика. Вона вкрита зверху протоплазматичним чохлом. Плазма сперматид розподіляється переважно у голівці, шийці і тілі спермія і завершує утворення сперміїв з характерною для них будовою – наявністю голівки, шийки, тіла і хвостика. Сформовані спермії розріджують за допомогою ферменту гіалуронідази протоплазматичний виріст клітини і виходять у просвіт каналика.
При розгляді під мікроскопом зрізу через звивистий каналик сім’яника можна по-бачити, що сперміогенез має хвилеподібний характер: впродовж звивистого каналика спостерігаються певні послідовні групи сперматогенного епітелію постійного клітин-ногоскладу на різних стадіях дозрівання. Повна серія цитологічних змін між двома од-наковими групами сперматогенних клітин у одній ділянці звивистого каналика назива-ється циклом сперматогенного епітелію. Різні автори ділять цей цикл на 8–12 стадій.
Тривалість циклу сперматогенного епітелію (стадія дозрівання, від поділу спер-матоцитів 2-го порядку до виходу сперміїв у просвіт сім’яних канатиків) складає у барана 10,4 дня, кнура – 8, бугая – 13,51, людини – 16 днів. Тобто, найінтенсивніше він перебігає у кнура.
Сформовані спермії поступають у просвіт покручених каналиків і переміщаються по них у прямі каналики, сітку сім’яника, сперміовиносні каналики і нарешті – у ка-нал придатка сім’яника.
Просуваються спермії по каналиках під тиском нових порцій сформованих гамет, а також під впливом скорочення еластичних волокон стінок сім’яних каналиків.
Застосовуючи рентгенівське опромінювання та мічені попередники ДНК, вдалося встановити тривалість часуміж першим поділом сперматогоній і появою сформованих сперміїв у придатку сім’яника, тобто “тривалість сперміогенезу” у різних видів тва-рин. Вона складає у бугая 50–55 днів, барана – 40–53, кнура – 35–40, півня – 24–27.
По придатку сім’яника спермії просуваються рухами війок сперміовиносних про-ток, перистальтичними скороченнями канала придатка, постійним виділенням секрету та сперміїв сім’яними каналиками, а також за рахунок еякуляції. У барана спермії про-ходять придаток за 10–14, а то і за 14–21 день, бугая – за 11, кнура – за 10–14 днів.
Розділ 2
При частому одержанні сперми від плідників проходження їх через придаток при-скорюється і в еякуляті зростає кількість неповноцінних сперміїв, хоча навіть при статевому виснаженні у придатку залишається великий запас сперміїв.
Добова спермопродукція у баранів складає біля 5 млрд, аукнура – 12–15 млрд. Вста-новлено тісний взаємозв’язок між масою сім’яників та їх репродуктивною здатністю.
Сперміогенез продовжується безперервно, але запаси сперміїв у придатку не пе-ревищують певної межі, оскільки вони регулюються особливим біологічним механіз-мом. Так, організм звільнюється від зайвих сперміїв шляхом ресорбції (фагоцитозу) або ж виділення з сечею. У бугаїв під час статевого спокою виявляли щодоби у добо-вій сечі від 125 до 3675 млн сперміїв, а при регулярному одержанні сперми – від 106 до 405 млн. У баранів ця цифра коливалась від окремих сперміїв і до 2477 млн.
Загальна довжина каналу придатка сім’яника складає у домашніх тварин від 30 до 86 м (у бугая – 30 м, кнура – 64, жеребця – 80 м). Тут міститься у барана від 100 до 220 млрд сперміїв, бугая – 50–75 млрд, кнура – 175–200 млрд.
У зв’язку з цим придаток сім’яника вважається своєрідним сперміосховищем. Тут вони зберігають свою живучість і запліднюючу здатність до двох місяців.
У голівці придатка сім’яника відбувається остаточне дозрівання сперміїв (за 6–8 днів). Проходження сперміями довгого шляху від місця утворення до місця збері-гання (а це біля 75 см довжини звивистого каналика, 14 см – по прямому каналику і приблизно 500 м загальної довжини сітки сім’яника) супроводжується дальшими змінами, що розпочалися ще з першого поділу сперматоцитів. Відбуваються не лише їх кількісні, а й якісні зміни ядра та протоплазми клітини.
У придатку сім’яника спермії набувають також від’ємного електричного заряду, який обумовлений електронегативним характером колоїдів поверхні клітин. Цей од-нойменний електричний заряд перешкоджає зіткненню і аглютинації сперміїв. При зменшенні електричного заряду сили відштовхування слабнуть і можливе зіткнення сперміїв і склеювання їх клейкою поверхнею. Такі спермії не можуть рухатися і при-йняти участь у процесі запліднення.
Проходження сперміїв вздовж каналу придатка супроводжується не тільки змі-нами їх фізіологічних властивостей, а й форми. Так життєздатність і запліднююча здатність сперміїв, добутих з хвоста придатка, значно вища, ніж сперміїв з голівки
Фізіологічні особливості статевих органів свійських тварин
придатка. Крім того, спермії голівки придатка мають на проксимальному кінці шийки так звану протоплазматичну краплю. З проходженням сперміїв крізь хвостову части-ну придатка крапля просувається на тіло спермія, а тоді спадає зовсім.
Накопичені у придатку сім’яника спермії поступово виводяться під час статевого акту чи одержання сперми на штучну вагіну. Якщо ж тривалий час плідник не вико-ристовується, то спермії старіють, відмирають і видаляються частково з сечею, част-ково піддаються резорбції, а на їх місце просуваються новосформовані.
Перебіг cперміогенезу, кількість та повноцінність утворених сперміїв, поряд з ге-нетичними факторами, у значній мірі визначаються умовами утримання, годівлі та догляду самців. Неповноцінна годівля, інтенсивна експлуатація, відсутність моціону, ожиріння, як і різноманітні захворювання і в першу чергу захворювання статевих ор-ганів негативно позначаються на сперміогенезі, кількості та якості виділюваних під час еякуляції сперміїв, їх живучості та запліднюючій здатності. У кінцевому рахунку вони можуть бути причиною неплідності.
Дата добавления: 2016-01-04; просмотров: 69; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!