Гаструляция у ланцетника
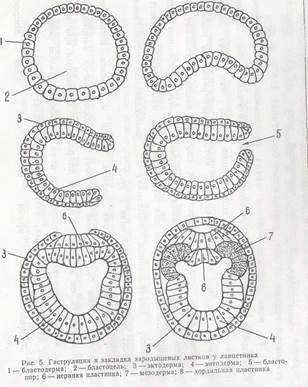
Гаструляция у ланцетника представляет удобную модель для изучения общих закономерностей этого процесса у типа хордовых. Этот процесс, как было показано ранее, у ланцетника, происходит по типу инвагинации. На ранней стадии гаструлы обнаруживается два зародышевых листка: эктодермальный, образованный из крыши бластулы, и энтодермальный— из дна бластулы. Гастроцель сообщается с внешней средой при помощи бластопора.
Наступает следующий этап развития зародыша — образование третьего зародышевого листка—мезодермы. Клетки дорзальной губы бластопора, быстро размножаясь, погружаются в составе энтодермы и на дорзальной ее стороне образуют хордальную пластинку— зачаток будущей хорды. Мелкие клетки вентральной и боковых губ бластопора также врастают в энтодермальный листок и располагаются в его составе по сторонам от хордальной пластинки.
Двуслойный зародыш приобретает вытянутую форму, имея в своем составе материал трех зародышевых листков. Материал мезодермы занимает дорзальную часть внутреннего листка. Из первичной эктодермы, расположенной над хордальной пластинкой, выделяется нервная пластинка. Последняя погружается и по длине зародыша образует нервный желобок. Нервный желобок, замыкаясь кверху, образует нервную трубку.
Хордальная пластинка выходит из состава первичного внутреннего листка, замыкается книзу и образует плотный клеточный тяж — хорду.
|
|
|
Одновременно мелкоклеточный материал, находившийся в первичном внутреннем листке по сторонам от хорды, выпячивается в дорзальном направлении в виде двух карманов. Вначале эти карманы открываются в гастроцель, а затем отделяются в виде двух замкнутых складок, расположенных вдоль гаструлы.
Так образуется третий зародышевый листок — мезодерма. После выделения хорды и мезодермы материал первичного внутренного листка смыкается, образуя энтодерму — внутренний зародышевый листок (рис. 5).
ГАСТРУЛЯЦИЯ у птиц
Гаструляция у птиц представляет довольно близкую модель процессам, имеющим место у высших хордовых, в том числе и у человека.
У птиц в результате дискоидального (меробластического) дробления образуется бластодиск. Он состоит из клеток неправильной формы, плотно прилегающих друг к другу и расположенных на огромной массе нераздробившегося желтка. Под влиянием ферментов клеток бластодиска часть желтка разжижается и образуется полость, заполненная жидкостью — бластоцель. Крыша этой бластулы представлена бластодиском, дно — нераздробившейся массой желтка. Бластоцель — подзародышевая полость.
Затем наступает гаструляция, состоящая из двух фаз. Первая фаза начинается до откладывания яйца и заключается в обособлении энтодермы путем деляминации клеток бластодиска. В результате возникает двуслойный зародыш. Верхний слой клеток дискобластулы приобретает призматическую форму и располагается в один правильный ряд. Нижний слой клеток дискобластулы сохраняет округлую или неправильную форму и лежит беспорядочно на желтке. Между клетками верхнего и нижнего слоя возникает полость — гастроцель.
|
|
|
Вторая фаза гаструляции начинается во второй половине первых суток инкубации. Бластодиск разрастается, в центре его выделяется зародышевый щиток, из которого в дальнейшем развивается тело зародыша. Окружающая зародышевый щиток часть дискобластулы представляет внезародышевый материал, в котором различают светлое и темное поле. Светлое поле расположено вокруг зародышевого щитка и состоит из клеток, отделившихся от желтка подзародышевой полостью в силу частичного использования желтка зародышем. Темное поле занимает периферию дискобластулы и состоит из клеток, плотно прилегающих к желтку и растущих по его поверхности. К этому моменту резко выражено разделение клеточного материала на зародышевую (зародышевый щиток) и внезародышевую (светлое и темное поле) часть.
|
|
|
В течение 24 часов насиживания или инкубации на зародышевом щитке в результате размножения клеток наблюдается перемещение их спереди назад по периферии зародышевого щитка. Оба потока клеток встречаются в центре у заднего конца зародышевого щитка, сливаются и перемещаются кпереди посредине зародышевого щитка. В результате образуется утолщенный тяж клеток, получивший название первичной полоски.
На переднем конце первичной полоски формируется утолщение— первичный узелок. В дальнейшем часть бластомеров, расположенных кпереди от первичного узелка, перемещается к последнему, подворачивается под эктодерму и формирует растущий кпереди между эктодермой и энтодермой головной или хордальный вырост — зачаток хорды.
Бластомеры задней половины зародышевого щитка смещаются к первичной полоске и через эту область погружаются под эктодерму, располагаясь в полости между эктодермой и энтодермой. Этот материал образует зачаток мезодермы — третьего зародышевого листка. Клетки хордального выроста размножаются и. перемещаются, занимая центральное положение между эктодермой
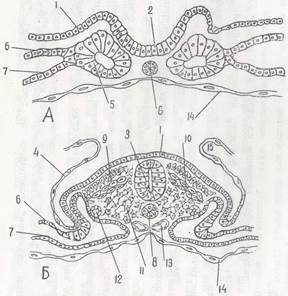
Рис.%6. Развитие комплекса осевых органов у птиц 1—эктодерма; 2 —нервный желобок; 3 — нервная трубка; 4 — амнион; 5 — сомиты; 6 — париетальный листок сплахнотома; 7 — висцеральный листок спланхнотома; 8 — хорда; 9 - дерматом; 10 — миотом; 11 — склеротом; 12 — нефротом; 13 — аорта; 14 — энтодерма; 15 — амниотическая складка и энтодермой в передней половине зародышевого щитка Вслед за этим клетки мезодермы также перемещаются в переднюю половину зародышевого щитка, располагаясь между эктодермой и энтодермой по бокам от хордального выроста. Это парахордальная мезодерма. Ее можно подразделить на дорзальную и вентральную.
|
|
|
По мере того как клетки первичной полоски уходят в закладки хорды и мезодермы на поверхности остается материал эктодермы и входящей в нее нервной пластинки. Таким образом, первичная полоска и головной узелок по своему значению соответствуют бластопору ланцетника, т. к. в этом месте проходит перемещение зародышевого материала, приводящее к образованию 3-зародышевых листков. Наиболее глубоко расположен зачаток кишечной энтодермы. Он непосредственно прилегает к поверхности желтка и находится во внутреннем слое дискобластулы.
К концу гаструляции в теле зародыша имеются все зачатки, из которых возникает типичный для хордовых комплекс осевых органов (рис. 6.).
НЕЙРУЛЯЦИЯ
Процесс, обеспечивающий переход зародыша от стадии гаструлы на стадию нейтрулы, получил название нейруляции. При этом, помимо нервной трубки, идет образование хорды и сомитов. Эта триада называется органами спинного комплекса.
В период нейруляции по средней линии на спинной стороне зародыша в результате размножения клеток образуется утолщение — нервная пластинка. Из этой же нейральной эктодермы формируются гребневидные складки — нервные валики, по бокам ограничивающие нервную пластинку. Нервные валики появляются сначала на переднем конце тела зародыша, а затем в средних и задних частях его.
Клетки нервной пластинки, размножаясь погружаются вовнутрь зародыша, образуя нервный желобок. Затем края этого желобка смыкаются и* возникает нервная трубка, содержащая полость — невроцель. Широкая передняя часть нервной трубки в дальнейшем даст структуры головного мозга, а более узкая туловищная часть — спинной мозг. Нервные валики приподнимаются над дорзальной поверхностью нервной трубки, сближаются друг с другом, формируют непарную ганглиозную пластинку. Эктодерма погружает под себя ганглиозную пластинку и нервную трубку, и в дальнейшем превращается в поверхностный хлой кожи--эпидермис. После смыкания кожной эктодермы ганглиозная пластинка располагается между ней и нервной трубкой и представляет собой тяж клеток, называемый нервным гребнем. Это временное скопление клеток, которое быстро распадается после возникновения.
Большинство клеток нервного гребня мигрирует по направлению к вентральной стороне зародыша.
Из клеток нервного гребня в дальнейшем образуются меланоциты — клетки, содержащие пигмент меланин, придающий коже окраску, а также, вероятно, клетки диффузной нейроэндокринной системы.
В самом начале нейруляции до появления нервной трубки по средней линии в передней половине зародышевого щитка из общего зачатка хордо-мезодермы обособляется хорда, по бокам которой располагается парахордальная мезодерма. Ее подразделяют на дорзальную и вентральную.
Дорзальная мезодерма в передне-заднем направлении -сегментируется на сомиты, в которых в дальнейшем выделяют три участка, получившие соответственно названия: дерматом, миотом и склеротом.
Вентральная мезодерма разделяется параллельно поверхности тела на два листка: париетальный, прилежащий к эктодерме, и висцеральный, прилежащий к энтодерме. Между этими листками появляется узкая щель — вторичная полость тела или целом.
Между дорзальной и вентральной мезодермой, объединяя их,
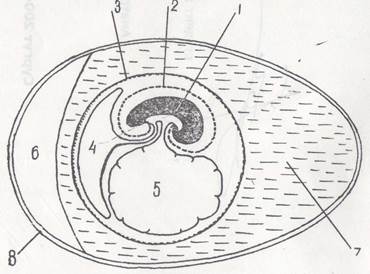
Рис. 7. Провизорные органы у птиц 1 — зародыш; 2 — амнион; 3 — серозная оболочка; 4 — аллантоис; 5 — желточный мешок; 6 — воздушная камера; 7 — белок; 8 — скорлупа
обособляются сегментные ножки или нефротомы, материал из которого в дальнейшем даст некоторые структуры мочеполовой системы. Общий план строения зародыша на поперечном срезе представлен на приводимой схеме (рис. 7).
ГИСТОГЕНЕЗ
Эмбриональный гистогенез — процесс возникновения тканей из малодифференцированных клеток эмбриональных зачатков. Эмбриональные зачатки (кожная эктодерма, кишечная энтодерма, нервная трубка, ганглиозная пластинка, миотом и др.) к концу нейруляции представлены сравнительно небольшими группами малодифференцированных клеток и не имеют тканевого строения. В отличие от тканей, в составе которых присутствуют специализированные клетки и могут быть структуры, не имеющие клеточного строения (симпласты, межклеточное вещество и др.), эмбриональные зачатки имеют вначале только клеточный состав.
Эмбриональная гистогенез — сложный процесс, который можно с известной степенью условности расчленить на 3 группы более простых состовляющих процессов. К первой группе можно отнести процессы изменения числа клеток эмбрионального зачатка за счет их размножения (а также гибели клеток). Ко второй группе — процессы изменения пространственного расположения клеток за счет их перемещения. К третьей группе — процессы дцфференциации клеток за счет изменения их структуры, размеров и_функции. В течение дифференциации клетки эмбрионального зачатка постепенно приобретают структурные и функциональные свойства, характерные для той или иной ткани. Параллельно с дифференциацией осуществляется повышение степени интегрировянности развивающихся тканей за счет взаимодействия клеток, составляющих эти ткани.
Перечисленные процессы необходимы для образования 4 групп тканей: эпителиальных, соединительных, мышечных, нервной. Частные закономерности формирования каждой из тканей будут рассмотрены в курсе общей гистологии. Здесь перечислим эмбриональные зачатки и развивающиеся из них ткани.

|

Дата добавления: 2016-01-05; просмотров: 114; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!