Транспорт через мембрану відбувається завдяки таким механізмам:
пасивного;
Первинно – активного; вторинно – активного.
Пасивний транспорт – відбувається завдяки спеціальним каналам без затрат енергії, завдяки дифузії.
Активний транспорт – відбувається завдяки спеціальним білковим структурам із затратами енергії. Первинно активний транспортує одну сполуку, вторинно – активний дві сполуки.
В одному напрямі рух (симпорт), в різних напрямах (антипорт).
Для пасивного транспорту є широкий набір білків переносників ( транспортних білків).
Воно проходить за рахунок дифузії, але високо специфічним структурам і набагато швидше.
Так транспортуються іони, аміно і органічні кислоти, моносахариди, нуклеотиди.
Найбільш типовим є транспорт іонів через спеціальні білки – переносники канали (пори).
Найбільш важливі канали: натрієві, калієві, кальцієві.
САМОСТІЙНА РОБОТА:
Канал має:
Водну пору, селективидний фільтр, і ворота.
Водна пора забезпечує проходження іонів зв’язаного з водою. Селективний фільтр має різні розміри для Na+ - 0,5 нм, для К +- 0,3 для Са+ - 0,65нм. Це дає можливість не всім проходити через канали. Ворота забезпечують положення «відкритого» і «закритого» яке залежить від: електричного заряду мембрани; спеціального рецептора який при взаємодії з лігандом їх відкриває.
| |

| |

| |

а) потенціал спокою б) деполяризація мембрани в) деполяризація мембрани,
|
|
|
що продовжується
m- активаційні «ворота»
h – інактивацій ні ворота
1- канал
2 – макромолекула білка
На рецептор діє ліганд, ворота відкриваються і іони проходять через канал.
Для активного транспорту використовуються насоси (помпи) Nа+, К+ АТФаза –натрієво, калієво – адинозинтрифосфатний насос. Насос, який забезпечує завдяки енергії молекули АТФ, викачування із клітини 3 молекул натрія, і закачування в клітину 2 молекул калія. Енергетично для роботи насоса використовується 24% всієї енергії клітини, в нервовій клітині 70%
Са2+АТФаза – кальцієво - адинозинтрифосфатний насос. Існує два види: один викачує із клітини кальцій в міжклітинну речовину; другий із цитоплазми (м’язевої) в саркоплазматичний ретікул.
Вторинно – активний транспорт наряду з викидом Na забезпечує надходження
других речовин. Наприклад вуглеводи і амінокислоти ентероцитами тонкої кишки.
Механізм транспорту речовин через мембрани:
|
|
|
В клітину можуть транспортуватися різні речовини за рахунок ендоцитоза піноцитоз і фагоцитоз.
Піноцитоз – поглинання речовин мембраною. На поверхні мембрани є рецептор (білок), який з’єднується поглинаючим білком. На внутрішній поверхні мембрани є актиномізіновий комплекс.
При з’єднанні рецептора з білком на поверхні мембрани під білком утворюється ямка край якої змикають скоротливі білки (актин –міозиновий комплекс).
 Білок Рецептор Актоміозиновий комплекс
Білок Рецептор Актоміозиновий комплекс

Фагоцитоз – поглинання другої клітини, бактерії, загинувши структур.
Рецептор мембрани з’єднується з білком клітини бактерії. Мембрана починає поступово обволікати клітину бактерії., поки бактерія не буде всередині клітини (фагоцита).
Макрофаг обволікає бактерію, цитоплазма якої у два рази більша за цитоплазму макрофага.
 Екзоцитоз - виділення із клітини ряд речовин, які вона синтезує. Органели утворюють везикули, мембрана якої зливається з мембраною клітин і речовина потрапляє за межі клітини.
Екзоцитоз - виділення із клітини ряд речовин, які вона синтезує. Органели утворюють везикули, мембрана якої зливається з мембраною клітин і речовина потрапляє за межі клітини.
Транспорт через мембрану йонів залежить від стану клітини, кількості утворення енергії, електричного заряду мембрани, рецепторів і т.д.
|
|
|
В стані фізіологічного спокою між клітиною і міжклітинною рідиною існує різниця потенціалів який називають мембранний потенціал спокою (МПС). Його величину можна виміряти якщо ввести у клітину мікроелектрод (скляний капіляр, заповнений розчином, який проводить струм).

Другий електрод помістити зовні мембрани. Всередині клітини заряд негативний, поза клітиною позитивний.
Для клітини скелетного м’яза МПС - 90мв. для гладеньких м’язів - 30мв., для нервової клітини -40 -90 мв., для секреторної клітини -20 мв.
Походження МПС.
В клітині і поза межами клітини неоднакова концентрація йонів.(ммоль/л)
| Іони | В клітині | Поза клітиною |
| Na+ К+ С2+ Сl- другі катіони другі аніони | 12 155 0.0001 4 - 155 | 145 4 2.4 120 5 7 |
Кількість катіонів (позитивно заряжені іони) і аніонів (негативно заряджені іони) всередині клітини рівна. Але мембрана клітини по різному прониклива для іонів. Аніони слабо проходять через мембрану. Завдяки йонним каналам катіони металів гарно проходять через мембрану. Проникливість її для К+ в 10-25 раз вища ніж Na+ в стані спокою. Тому кількість виходу К+ буде набагато більше ніж вступ до клітини Na+ , що утворює різницю потенціалів, яку можна зафіксувати. Всередині клітини із зменшенням К+ зміниться потенціал на негативний а поза клітиною внаслідок накопичення К+ стане позитивний.
|
|
|
Якщо вихід К+ з клітини і вхід Na+ в клітину буде так і продовжуватися, то з часом не буде різниці потенціалів. Але цього не допускає Na+ К+ - насос, який відкачує натрій з клітини і закачує в неї калій підтримуючи МПС. МПС відображає обмін речовин в клітині, утворення АТФ. При зміні обміну речовин та порушенні синтезу АТФ падає МПС.
Для того, щоб змінити МПС (мембранний потенціал спокою) потрібно клітину подразнити і визвати збудження.
Мінімальна сила подразника, яка визиває збудження називають пороговим.
Менша сила чим порогова змінює фізико – хімічні властивості тканини, але не визиває збудження називають підпороговою.
Подразник, в якого сила більша за порогову називають надпороговим.
В експериментах Лапін на м’язах і нервах застосовував в якості подразника постійний електричний струм. Мінімальна сила постійного струму, здатна викликати збудження (поріг подразнення) назвали реобазою.
Найменший час на протязі якого повинен діяти подразник величиною в одну
реобазу називають корисним часом. Тому збільшувати час дії подразника немає практичного значення. З збільшенням сили подразника зменшується мінімальний час дії його, але не безмежно. (дивись криву)
Тому ввели поняття хронаксія.
Хронаксія – це час на протязі якого діє струм у 2 реобази, щоб визвати збудження. Величину хронаксії і реобази використовують в медичній практиці.
Якщо у хворого пошкоджений нерв, то хронаксія його буде значно більша.
Дослідження 19 століття Л. Гальвані показали, що живі тканини можуть бути джерелом електричного потенціалу. Через 100 років нідерландський фізіолог Єйнтховен винайшов струмний гальванометр, що даломожливістьзаписати на папері електричні потенціали діяльності серця. Так в медицину ввійшла електрокардіографія, енцефалографія.
Винайдення мікроелектродів, дало можливість записувати електричні потенціали на мембрані клітин, в середині клітин, мозку і т.д.
Німецький фізіолог Дюбуа –Реймон запропонував метод електричного подразнення тканин і клітин, що дало практичній медицині електростимулятори серця, м’язів, нервової тканини.
При дії на тканини різними хвилями (ультразвук, лазер, теплові) можна спеціальними датчиками зафіксувати зміни в параметрі тканин, що дало можливість винайти методи дослідження ультразвукові, реографи і т.д.
Сучасний прорив в дослідженні організму його структури, функції. діагностики хвороб дала магнітно – резонансна комп’ютерна томографія.
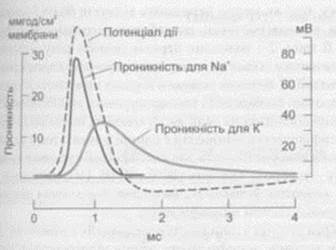
Якщо на нерв, або м’яз нанести подразник порогової сили виникає зміна потенціалу. Потенціал дії реєструється на осцилографі за допомогою мікроелектродів. ПД і є збудження.
Крива ПД (потенціалу дії) має фази.

1 –фаза наростання деполяризації
2 – критичний рівень деполяризації (поріг)
3 - овершут (переліт)
4 - реполяризація (зниження)
5 - слідова деполяризація
6 -слідова гіперполяризація
При дії подразнення потенціал зменшується до – 55 мв. Це називається критичний рівень деполяризація (поріг), після чого йде швидке зменшення до 0 і +30,+35 мв. (овершут) – переліт потенціалу. Далі наступає швидка реполяризація, яка сповільнюється (слідові деполяризація). В подальшому переходить у невелику слідові гіперполяризацію.
В основі виникнення ПД лежать іонні механізми.
При подразненні клітини відкриваються натрієві ворота і натрій лавиноподібно починає надходити в клітину міняючи потенціал, що викликає деполяризацію мембрани. Натрієві канали швидко активізуються, а калієві повільно.
Тому вихід К+ в період деполяризації не перешкоджає процесу деполяризації мембрани. Внаслідок деполяризації відкриваються К+ канали, а повільно закриваються Na+ і наступає реполяризація мембрани.
Збільшення Na+ в клітині (в період деполяризації), вихід К+ з клітини (в період реполіризації) включає Na+ К+ насоси,які відновлюють вміст йонів в клітині.
 |
Потенціал дії через іонні механізми забезпечує збудження.
Виникнення ПД супроводжується зміною збудливості:
1) Начальне підвищення збудливості – до критичного рівня деполяризації.
2) Рефрактерний період, який поділяється на абсолютну рефрактерність (АР) при якій навіть над порогові подразнення не викликають збудження
Цей період часу наступає з моменту критичної деполяризації до точки
проходження однієї третини реполяризації: Абсолютна незбудливість
пояснюється неможливістю збільшення натрієвої проникливості мембран.
Відносно рефрактерний період триває від точки закінчення АРТ до початку слідової деполяризації. В цей період подразник більшої сили чим
подразник, який викликав подразнення може викликати збудження.
 |
3) Супернормальний період – період по часу відповідає слідовій деполяризації:
В цей період тканина відповідає збудженням на субпороговий подразник.
Збудливість підвищена.
4) Субнормальний період – період по часу відповідає слідовій гіперполяризації.
Збудливість незначно знижена.
Якщо на тканину подають катодний струм який спричинює деполяризацію рівня 7мв., то вона не викликає ПД. Там просто пасивно додаються негативні заряди. Катодний струм що спричиняє деполяризацію 7-15 мв. також не викликає ПД. Він викликає лише часткове збільшення проникливості Na+ каналів. Такі зміни проникливості називають локальною відповіддю.
У разі деполяризації 15мв. сили деполяризації достатні щоб переважити сили реполяризації і викликає потенціал дії. Точка при якій відбувається різке сходження потенціалу називають критичним рівнем деполяризації. Виникнення подразника підпорогового викликає локальну відповідь, а ПД виникає коли подразник пороговий або надпороговий. Отже потенціал дії підлягає закону «все або нічого»
Подразнення звичайно подають з катода, оскільки катодні подразнення мають деполяризований вплив. Анодні струми пригнічують утворення імпульсу внаслідок гіперполяризації. Однак припинення анодного струму може спричинити ПД і виникає збудження. Цей закон, який установив Пфлюгер у 1859 р. називають полярний закон подразнення . «При подразненні тканини постійним струмом збудження виникає під катодом у момент вмикання постійного струму, під анодом в момент вимикання».
При подачі постійного струму на електроди, які розміщені на поверхні клітин, потрібно давати більшу силу струму. Тому що струм проходить не тільки по мембранах, але й по міжклітинних щілинах. При розміщенні одного електрода всередині клітини сила струму подразнює тільки мембрану ПД який виникає як відповідь на подразнення розповсюджується по нервовому волокну.
Нервові волокна є безмієлінові та мієлінові. Нервові волокна безмієлінові покриті тільки оболонкою навколо осьового циліндра нервового відростка. При подразненні безмієлінового волокна позитивно перезаряджені мембрани попереду і позаду ПД переходить у ділянку, де містяться негативні заряди. Завдяки переміщенню позитивних зарядів зменшується полярність мембран попереду, така електротонічна деполяризація спричиняє локальну відповідь, критичний рівень і збудження поширюється, що відповідно далі попереду електротонічно деполяризує мембрану. Завдяки такій почерговості збудження рівномірно поширюється до його кінця. Збудження в експерименті поширюється в різні кінці нервового волокна, а збудження в організмі тільки в одному напрямі.
Мієлінові оболонки мають спеціальну мієлінову оболонку, яка ізолює нервове волокно. Крім того у мієлінових волокнах є перетяжка Ранв’є. Натрієві канали мають практично тільки у перетяжках Ранв’є і тому деполяризація виникає між двома перетяжками стрибкоподібно (сальтаторно), і ому у 50 раз швидше чим самі швидкі безміалінові.

Швидкість розповсюдження збудження по нервовому волокну залежить від мієлінової оболонки, товщини нервового волокна. Чим більший поперечний зріз волокна тим більша швидкість.
Швидкість збудження по мієліновому волокну від 70-120 м/с, по бізмієліновому 0,5 -2 м/с.
Дата добавления: 2018-10-27; просмотров: 596; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!