Уровни упаковки ДНК. Строение хромосом
Упаковка ДНК включает несколько уровней (рис.11).
1) Первый уровень заключается в образовании нуклеосомной нити. Нуклеосома состоит из октамера белков-гистонов, обвитого 2 витками ДНК. Октамер (греч. octo – восемь, meros - часть) состоит из 8 глобулярных белков-гистонов (парные Н2А, Н2В, Н3, Н4). Нуклеосомы разделены короткими участками линкерной ДНК. С октамером и частично с участком линкера связывается гистон H1. Цепь нуклеосом называется нуклеосомной нитью. Она имеет диаметр ~10нм. ДНК укорачивается в 6-7 раз.
2) Второй уровень упаковки – образование хроматиновой фибриллы. При этом нуклеосомная нить закручивается в спираль диаметром 30 нм. ДНК укорачивается в 40 раз. В дальнейшей упаковке хроматина ведущую роль играют негистоновые белки.
3) Третий уровень упаковки ДНК – хромомерный. Хроматиновые фибриллы образуют петли – хромомеры, диаметром около 100-150 нм. ДНК укорачивается в 600 раз. Нитчатая хроматиновая структура со средней толщиной 100-200 нм получила название хромонемы. Хромонему можно наблюдать на стадии начальной конденсации хромосом в профазе митоза и при деконденсации хромосом в телофазе.
4) Четвертый уровень - хромосомный. Заключается в более плотной упаковке петель и образовании «конденсированных участков хромосом» диаметром около 700 нм. Максимальной степени конденсации хромосомы достигают в метафазе митоза/меойза.
Строение хромосом (рис.11).
Хромосомы на стадии метафазы образованы двумя хроматидами, объединенными в области первичной перетяжки – центромеры. Каждая хроматида представляет собой дочернюю ДНК. В зонах центромер расположены белковые структуры – кинетохоры, они являются местом прикрепления нитей веретена деления. Участки, на которые центромера делит хромосому, называются плечи, концы хромосомы – это теломеры.
|
|
|
Некоторые хромосомы имеют вторичные перетяжки (ядрышковые организаторы), они содержат гены, кодирующие рРНК. Часть хромосомы, отделенная вторичной перетяжкой, называется спутник или саттелит.
По положению центромеры хромосомы делятся на 3 вида (рис.12):
1) Метацентрические – хромосомы, которые имеют равные плечи
2) Субметацентрические – хромосомы с неравными плечами
3) Акроцентрические – хромосомы, у которых одно плечо практически отсутствует.
|

|





|

|
|
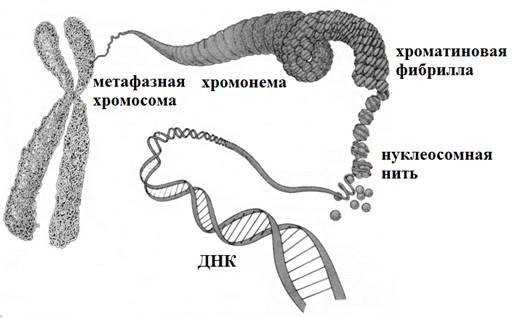
Рис. 11. Упаковка ДНК. Строение метафазной хромосомы
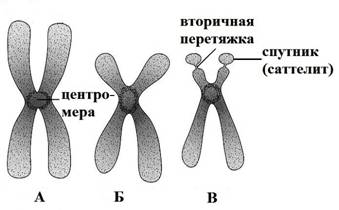
Рис. 12. Форма хромосом: А – метацентрическая хромосома,
Б – субметацентрическая хромосома, В – акроцентрическая хромосома
Метаболизм
Метаболизм – обмен веществ и энергии. Включает:
|
|
|
1. Пластический обмен (ассимиляция или анаболизм) – синтез органических веществ с затратой энергии.
2. Энергетический обмен (диссимиляция или катаболизм) – распад органических веществ с освобождением энергии.
Синтез белка
1. Транскрипция -синтез иРНК на матрице ДНК.
В основе транскрипции лежит принцип комплементарности азотистых оснований ДНК и РНК. Информационная РНК строится в направлении от 5′ -к 3′-концу, антипараллельно матричной цепи ДНК. В качестве матрицы используется лишь одна из двух цепей ДНК – она называется матричной (или значащей, или кодогенной).
Транскрипция происходит на определенном участке ДНК – транскриптоне. Транскриптон ограничен двумя последовательностями, которые называются промотором (зона начала транскрипции) и терминатором (зона остановки транскрипции). Транскриптон эукариот соответствует одному гену. Транскриптон бактерий называется оперон и включает несколько структурных генов. Транскрипция осуществляется с помощью фермента – РНК-полимеразы.
Первичные РНК, которые образовались в результате транскрипции, представляют собой функционально неактивные молекулы. Созревание РНК называется процессингом. Одним из механизмов процессинга мРНК является сплайсинг. Сплайсинг – это вырезание из пре-мРНК некодирующих областей (интронов) и сшивание кодирующих структуру белка участков (экзонов).
|
|
|
Зрелая мРНК через ядерные поры выходит в цитоплазму, где осуществляется трансляция (рис.13).
2. Трансляция– процесс перевода генетической информации с последовательности нуклеотидов мРНК в последовательность аминокислот в молекуле полипептида. Трансляция осуществляется согласно правилам генетического кода:
1. Код – триплетный, т. е. одну аминокислоту определяет три нуклеотида.
2. Код – однозначный (специфичный): каждый кодон обозначает только одну аминокислоту.
3. Код – вырожденный, т. е. одной аминокислоте может соответствовать более одного кодона.
4. Код – непрерывный, т. е. отсутствуют сигналы, показывающие конец одного кодона и начало следующего.
5. Код не перекрывается, т. е. один нуклеотид не может одновременно входить в два соседних триплета.
6. Генетический код содержит триплеты, обозначающие начало и окончание синтеза белка. АУГ – инициирующий кодон (кодирует метионин). УАА, УАГ, УГА – терминирующие кодоны, которые не кодируют ни одну из известных аминокислот и сигнализируют об окончании синтеза белка.
|
|
|
7. Генетический код универсален, т. е. одинаков у животных, растений, многих бактерий.
Подготовительные стадии трансляции включают:
· активацию аминокислот;
· присоединение аминокислот к тРНК.
Обе стадии осуществляются с помощью фермента – аминоацил-тРНК-синтетазы (кодазы). Комплекс тРНК и аминокислоты называется аминоацил-тРНК (аа-тРНК).
Этапы трансляции
1. Инициация трансляции – начало синтеза полипептидной цепи, заключается в сборке активной рибосомы. Инициация трансляции начинается со связывания мРНК в области старт-кодона с малой субъединицей рибосомы. Далее происходит связывание первой аа-тРНК и присоединение большой субъединицы рибосомы.
«Собранная» рибосома имеет 2 активных центра:
· пептидильный центр (П-центр). В начале трансляции с пептидильным центром связывается первая инициирующая аа-тРНК. На последующих стадиях трансляции в пептидильном центре находится тРНК, содержащая уже синтезированный пептид.
· аминоацильный центр (А-центр) – место связывания очередной аа-тРНК. А- и П- центры расположены как на большой, так и на малой субчастицах рибосомы.
2. Элонгация трансляции – самый продолжительный этап синтеза белка, в ходе которого происходит удлинение полипептидной цепи за счет присоединения аминокислот. Начинается с момента образования первой пептидной связи и заканчивается после включения в полипептидную цепь последней аминокислоты. Элонгация трансляции имеет циклический характер. Цикл элонгации включает 3 стадии:
а) связывание аа-тРНК с А- центром рибосомы. Связывается аа-тРНК, чей антикодон комплементарен кодону мРНК.
б) образование пептидной связи между аминокислотами. Первая аминокислота переносится на вновь поступившую аминокислоту. Первая тРНК становится свободной.
в) транслокация – перемещение рибосомы по мРНК на один кодон. Свободная тРНК вытесняется из рибосомы, тРНК с дипептидом оказывается в П-центре, А-центр становится свободным. В А-центр приходит очередная аа-тРНК. Цикл повторяется.
3. Терминация трансляции – окончание синтеза белка. Сигналом об окончании трансляции служит появление в рибосоме одного из стоп- кодонов мРНК: УАА, УАГ или УГА. С терминирующим кодоном, который находится в А-центре, взаимодействуют особые белки. В результате нарушается связь полипептида с тРНК. Синтезированный белок отделяется от рибосомы, одновременно отделяются тРНК и мРНК, а рибосома диссоцирует на субъединицы.

Рис. 13. Синтез белка в эукариотичской клетке
Фотосинтез
Фотосинтез – синтез органических веществ (глюкозы) из неорганических (углекислого газа и воды) с использованием солнечного света. Осуществляется в хлоропластах. Важную роль в фотосинтезе играют пигменты: хлорофиллы (у высших растений а и в), каротиноиды. Основной пигмент – хлорофилл а. Совокупность молекул пигментов с белками-переносчиками электронов называется фотосистемой (или квантосомой). Выделяют две фотосистемы:
ФСI в качестве реакционного центра содержит хлорофилл, поглощающий свет с длиной волны 700 нм.
ФСII в качестве реакционного центра содержит хлорофилл, поглощающий свет с длиной волны 680 нм.
Фотосинтез включает две фазы:
1. Световая фаза заключается в преобразовании энергии квантов света в энергию химических связей АТФ. Также называется фотосинтетическим фосфорилированием. Протекает на внутренних мембранах хлоропласта.
Под действием кванта света электрон выбивается из ФСII и поступает в электронно-транспортную цепь (ЭТЦ). ЭТЦ состоит из белков-переносчиков, встроенных в мембрану тилакоида. Электроны, идущие по ЭТЦ, не выходят за пределы мембраны тилакоида, а передаются внутри мембраны от ФСII к ФСI. Одним из первых электрон принимает пластохинон. Наряду с электронами, пластохинон способен переносить и протоны. Он работает как «водородная помпа»: взяв электрон, пластохинон перекачивает протоны с наружной поверхности мембраны тилакоида на внутреннюю, а электрон передает далее по цепи.
Место выбитых электронов в ФСII занимают электроны, образующиеся при фотолизе воды. В результате фотолиза воды кроме электронов образуется кислород, который выделяется в атмосферу, и протоны, которые накапливаются внутри тилакоида.
Таким образом, в результате фотолиза воды и работы пластохинона количество протонов в строме убывает, а внутри тилакоида увеличивается, т.е. на мембране тилакоида создаётся градиент концентрации протонов. При достижении критической разности потенциалов (200мВ) протоны устремляются по протонному каналу в ферменте АТФ-синтетаза наружу. Пропуская через себя поток протонов, АТФ-синтетаза синтезирует АТФ из АДФ и фосфата, т.е. осуществляет фосфорилирование.
Квант света также выбивает электрон из ФСI. Выбитый электрон ФСI подхватывается переносчиком ферредоксином и выносится за пределы мембраны тилакоида. Электроны присоединяются к НАДФ+, восстанавливая его и одновременно присоединяя протоны, образуется НАДФ·Н2. Процесс переноса электронов от ФСII к ФСI, а затем к НАДФ+ получил название нециклического фосфорилирования (рис.14).
Наряду с нециклическим в мембранах хлоропластов происходит и циклический транспорт электронов, который осуществляется с участием только ФСI и ЭТЦ. В этом случае протонный градиент создаётся только за счёт работы пластохинона. Происходит синтез АТФ без фотолиза воды и восстановления НАДФ. Такой транспорт электронов преобладает в стрессовых условиях для растения (засуха, высокая температура, инфицирование), когда необходима срочная наработка энергии в виде АТФ.
Таким образом, в результате световой стадии фотосинтеза происходит:
• синтез АТФ;
• восстановление НАДФ·Н2;
• выделение О2 при фотолизе воды.
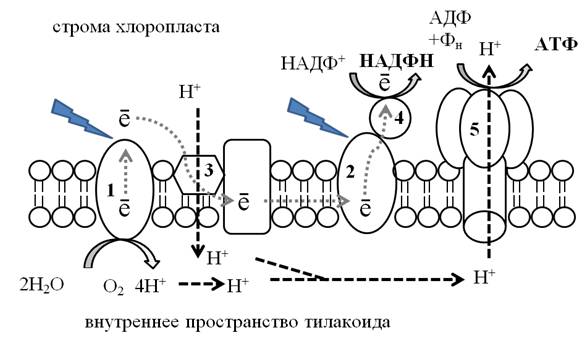
Рис. 14. Световая фаза фотосинтеза: 1 –фотосистема I, 2 – фотосистема II,
3 – пластохинон, 4 – ферредоксин, 5 – АТФ-синтетаза
2. Темновая фаза. Осуществляется в строме хлоропласта. На этом этапе фотосинтеза происходит восстановление углекислого газа до углеводов с использованием НАДФ·Н2 и АТФ, образованных в световую стадию. Процесс происходит циклично и называется циклом Кальвина. Цикл Кальвина можно разделить на три этапа:
а) Карбоксилирование. Поступающий в хлоропласт углекислый газ вначале присоединяется к акцептору - рибулозодифосфату. Образуется промежуточное нестойкое шестиуглеродное соединение, которое затем распадается на две молекулы трехуглеродной фосфоглицериновой кислоты (3-ФГК).
б) Восстановление. 3-ФГК восстанавливается до 3-фосфоглицеринового альдегида (3-ФГА). При этом используется АТФ и НАДФ·Н2, синтезированные в световой фазе фотосинтеза.
в) Регенерация. На этом этапе вновь образуется акцептор углекислого газа – рибулозодифосфат (РДФ). При фиксации шести молекул углекислого газа образуется 12 молекул 3-ФГА. 10 из них используется для регенерации 5 молекул РДФ, а из двух оставшихся триоз образуется гексоза (глюкоза).
Значение фотосинтеза.
1. За счёт фотосинтеза образуется органическое вещество (160-200 млрд. т. в год). Образующееся в процессе фотосинтеза органическое вещество служит пищей для животных и человека и сырьём для производственной деятельности.
2. В процессе фотосинтеза аккумулируется энергия солнечного света (космическая роль растений, К.А. Тимирязев).
3. В ходе фотосинтеза выделяется кислород (100-150 млрд. т. в год), необходимый для дыхания большинства организмов на Земле. Из кислорода образуется озон, который защищает все живое от ультрафиолетовых лучей.
4. В результате фотосинтеза растения поглощают углекислый газ, который вовлекается в природный кругооборот. Фотосинтез регулирует концентрацию углекислого газа в атмосфере и предотвращает парниковый эффект.
Энергетический обмен
Энергетический обмен заключается в расщеплении сложных органических веществ с выделением энергии, часть которой запасается в АТФ. Осуществляется в три этапа.
I. Подготовительный. Сложные органические вещества под действием ферментов распадаются на простые:
• белки до аминокислот,
• жиры до глицерина и жирных кислот,
• полисахариды до моносахаридов (крахмал до глюкозы).
Осуществляется в желудочно-кишечном тракте и лизосомах, выделяющаяся энергия рассеивается в виде тепла.
II. Бескислородный (анаэробный этап или гликолиз). Осуществляется в гиалоплазме, не требует присутствия кислорода. При распаде одной молекулы глюкозы образуется: две молекулы пировиноградной кислоты (ПВК), 4 атома водорода (они связываются с переносчиком водорода НАД+, образуя 2НАД·Н2) и выделяется энергия. Около 40% этой энергии аккумулируется в двух молекулах АТФ, остальная энергия (60%) рассеивается в виде тепла.
Дальнейшая судьба ПВК зависит от наличия кислорода в клетке. Если кислород есть, то ПВК окончательно окисляется в митохондриях до углекислого газа и воды. Если кислорода нет, то ПВК превращается либо в этиловый спирт (спиртовое брожение у дрожжей), либо в молочную кислоту (анаэробный путь в клетках животных, молочнокислых бактерий).
III. Кислородный (аэробный). Осуществляется в митохондриях в присутствии кислорода.
а) ПВК поступает в матрикс митохондрии, где декарбоксилируется (отщепляется углекислый газ) и дегидрируется (отщепляется водород, который переносится на акцептор, образуется НАД·Н2). Во время этих реакций ПВК (3С) превращается в ацетил (2С). Ацетил соединяется с коферментом А, образуя ацетил~КоА.
б) Ацетильная группа (2С) после гидролиза ацетил~КоА включается в цикл Кребса. Ацетил (2С) присоединяется к щавелевоуксусной кислоте (4С), в результате образуется лимонная кислота (6С). Энергию для этой связи поставляет макроэргическая связь ацетил~КоА. Далее следует цикл реакций, в результате которых ЩУК регенерируется, образуется 1 АТФ, 2 молекулы углекислого газа и 4 пары атомов водорода, которые переносятся на НАД или ФАД, а затем поступают в дыхательную цепь.
в) Дыхательная цепь представляет собой ряд белковых комплексов, встроенных во внутреннюю мембрану митохондрий (рис.15). Функционирование дыхательной цепи осуществляется в следующей последовательности:
• атомы водорода поступает на внутреннюю мембрану митохондрии в составе НАДН или ФАДН2, где окисляются (отдают электроны и превращаются в протоны);
• электроны передаются по ферментам дыхательной цепи к кислороду, который соединяется с протонами матрикса и образует воду;
• часть энергии электронов используется комплексами дыхательной цепи для перемещения протонов через внутреннюю мембрану в межмембранное пространство;
• в результате формируется градиент ионов водорода между наружной и внутренней поверхностями мембраны крист. Такой градиент получил название электрохимический (или протонный) градиент;
• при достижении определенного значения протонного градиента (соответствует разности потенциалов 220 мВ) АТФ-синтетаза начинает транспортировать протоны обратно в матрикс. Проходя через АТФ-синтетазу протоны теряют свою энергию. Большая часть этой энергии идет на синтез АТФ из АДФ и неорганического фосфата.
Совокупность процессов, происходящих на внутренней мембране митохондрий, получила название окислительное фосфорилирование. Окисление восстановленных переносчиков (НАДН и ФАДН2) ферментами дыхательной цепи сопряжено с синтезом АТФ.
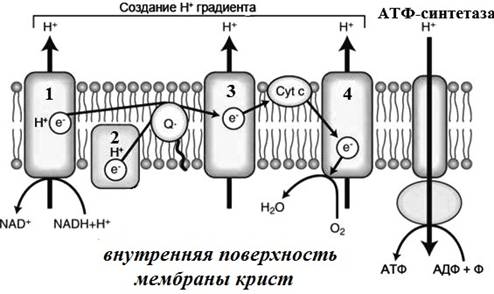
Рис. 15. Окислительное фосфорилирование на мембране крист:
1-4 – белковые комплексы дыхательной цепи
Таким образом, расщепление одной молекулы глюкозы до углекислого газа и воды обеспечивает синтез 38 молекул АТФ: на II этапе (гликолиз) синтезируется 2 АТФ, а на III этапе (кислородном) – 36 АТФ.
Жизненный цикл клетки
Жизненный цикл клетки(клеточный цикл) – период существования клетки от её образования до собственного деления или смерти. Жизненный цикл клетки состоит из митоза и промежуточного периода – интерфазы. Интерфаза занимает около 90% жизненного цикла клетки.
Периоды интерфазы:
1. Пресинтетический или постмитотический период (G1) начинается после завершения митоза и длится от нескольких часов до нескольких дней. Он характеризуется интенсивным синтезом РНК и белка, увеличением количества органоидов и активным ростом клетки.
2. Синтетический период (S) характеризуется удвоением молекул ДНК (репликация), синтезом белков-гистонов и удвоением центриолей.
Репликация (редупликация) ДНК - удвоение молекул ДНК. Репликация является матричным процессом, т.к. каждая дочерняя цепь ДНК строится, используя в качестве матицы родительскую ДНК. В основе репликации – принцип комплементарности (А-Т, Г-Ц). При этом матрицами служат обе цепи родительской ДНК. Репликацию можно назвать полуконсервативным процессом, т.к. после завершении репликации исходные молекулы ДНК оказываются наполовину обновленными. В каждой из дочерних молекул одна цепь – родительская, а вторая – вновь синтезированная. Удлинение цепи ДНК всегда происходит в направлении от 5′- к 3′-концу, т. е. очередной нуклеотид присоединяется к 3′-концу растущей цепи. Синтезируемая цепь антипараллельна матричной цепи.
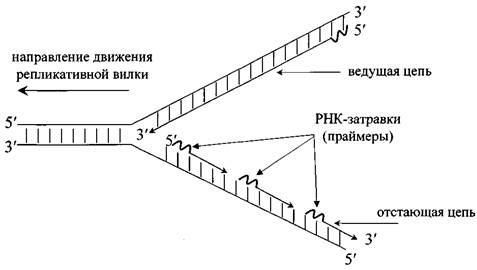 Для эукариотических клеток характерно наличие множества точек начала репликации. Репликация распространяется в обе стороны от каждой точки начала репликации. При этом образуются две репликативные вилки, которые движутся в противоположных направлениях. Одна дочерняя цепь синтезируется непрерывно – она называется ведущей или лидирующей. Вторая цепь ДНК синтезируется короткими фрагментами - фрагментами Оказаки. Эта цепь называется отстающей или запаздывающей (рис.16).
Для эукариотических клеток характерно наличие множества точек начала репликации. Репликация распространяется в обе стороны от каждой точки начала репликации. При этом образуются две репликативные вилки, которые движутся в противоположных направлениях. Одна дочерняя цепь синтезируется непрерывно – она называется ведущей или лидирующей. Вторая цепь ДНК синтезируется короткими фрагментами - фрагментами Оказаки. Эта цепь называется отстающей или запаздывающей (рис.16).
Рис. 16. Репликативная вилка
Основной фермент репликации – ДНК-полимераза. Однако ДНК-полимераза не может начинать процесс «с нуля», поэтому начинает синтез фрагментов дочерней ДНК другой фермент - ДНК-праймаза. ДНК-праймаза синтезирует РНК-затравку (праймер) – короткую последовательность из РНК-нуклеотидов. После возникновения 3′- конца растущего полинуклеотида процесс продолжает ДНК-полимераза. На завершающем этапе репликации РНК-затравки вырезаются, а отдельные фрагменты ДНК сшиваются лигазой.
3. Постсинтетический (или премитотический, G2) период включает подготовку клетки к делению: запасание энергии в АТФ, созревание центриолей, синтез иРНК и белков (в первую очередь тубулина).
Митоз
Митоз(греч. mitos - нить) – непрямое деление клетки. Митоз подразделяют на 4 фазы: профаза, метафаза, анафаза, телофаза.
1. Профаза. Хроматин конденсируется в хромосомы. Каждая хромосома состоит из двух сестринских хроматид. Транскрипция полностью прекращается. Исчезают ядрышки, ядерная оболочка фрагментируется. Центросомы начинают расходиться друг от друга на некоторое расстояние. В конце профазы начинается образование веретена деления. Набор хромосом 2n4c.
2. Метафаза. Хромосомы, состоящие из двух хроматид, располагаются по экватору клетки. К центромерам хромосом прикрепляются микротрубочки веретена деления. Контакт между хроматидами сохраняется в обрасти центромеры. Набор хромосом 2n4c.
3. Анафаза начинается с разъединения сразу всех хромосом в центромерных участках. Сестринские хромосомы, состоящие из одной хроматиды, синхронно расходятся к противоположным полюсам клетки. Анафаза – самая короткая стадия митоза. У каждого полюса набор хромосом 2n2c, а в целом в клетке 4n4c.
4. Телофаза. Хромосомы деконденсируются, ядерная оболочка восстанавливается, появляется ядрышко. Разрушается веретено деления. Телофаза заканчивается разделением исходной клетки на две дочерние – происходит цитокинез. Образуются 2 дочерние клетки с набором 2n2с.
Значение митоза. Митоз обеспечивает:
· рост и развитие организма;
· регенерацию;
· бесполое размножение.
Митоз – способ деления соматических клеток. Из одной диплоидной клетки образуются две равнозначные диплоидные клетки.
Мейоз
Мейоз (от греч. meiosis – уменьшение) – способ деления клеток, в результате которого вдвое уменьшается число хромосом. Из одной диплоидной клетки образуется четыре гаплоидных.
Мейоз состоит из двух делений.
Дата добавления: 2018-05-13; просмотров: 12663; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!