Классификация экзокринных желез
Эозинофильный гранулоцит. Электронные микрофотографии. 1. На левом снимке - эозинофильный лейкоцит целиком, а на правом - одна из его специфических (эозинофильных) гранул при очень большом увеличении - на продольном и на поперечном срезах. 2. а) На первом снимке хорошо видны: двудольчатое ядро (т.е. ядро, разделённое на 2 сегмента, между которыми остаётся узкая перемычка); митохондрии (например, справа от ядерной перемычки), а также многочисленные гранулы с плотными тельцами в середине. б) Последние - это специфические для эозинофиловоксифильные гранулы. 3. а) Данные гранулы имеют цилиндрическую форму и содержит кристаллоид - плотную пластинчатую структуру тоже цилиндрической формы. б) Эта структура образована т.н. щелочным белком. в) В целом же оксифильные гранулы эозинофилов - эторазновидность пероксисом: в них, видимо, происходит окислительное дезактивирование гистамина и серотонина, сопровождаемое образованием Н2О2. г) Кроме того, в этих гранулах содержится фермент пероксидаза, которая катализирует окисление веществ пероксидом водорода (Н2О2). 4. В итоге, эозинофилы обладают, как минимум, двумя видами активности: антигистаминной (т.е. противовоспалительной и антиаллергической), а также противопаразитарной (видимо, за счёт щелочного белка). Нейтрофилы — это наиболее многочисленная группа клеток белой крови (лейкоциты), основной задачей которой является уничтожение болезнетворных бактерий в крови и тканях. Нейтрофилы — это клетки-камикадзе. Встречаясь с бактерией в организме, нейтрофил поглощает её (фагоцитоз), расщепляет внутри себя (лизис), и далее погибает. Нейтрофилы имеют шесть стадии созревания, на двух из которых клетки находятся в крови в норме, и на двух из которых иногда могут появляться в крови, в случае "тяжелого" течения заболевания. Одновременно в организме находятся все шесть групп, но нам интересны только последние четыре, а также их соотношение между собой, которое является одним из важнейших диагностических критериев доступных в общем анализе крови, известным как сдвиг лейкоцитарной формулы. Давайте рассмотрим эти подгруппы нейтрофилов по степени созревания. Миелоциты -> Метамиелоциты -> Палочкоядерные нейтрофилы -> Сегментоядерные нейтрофилы. Старшие формы клеток первыми берут на себя удар при появлении болезнетворного агента, и, как мы сказали ранее, погибают. В большинстве случаев их работы достаточно для подавления болезни, но если инфекция очень тяжелая, то в «бой за кровь» могут вступать и средние клетки, и даже незрелые миело- и метамиелоциты, которые в норме проходят полный цикл созревания в костном мозге и у здоровых людей вообще отсутствуют в крови. Т.е. при тяжелом заболевании сегментоядерные нейтрофилы погибают первыми, и если организм не успеет синтезировать новые клетки, то в анализе крови будет мало тех клеток, которые в порядке созревания идут последними (справа) и больше молодых (в записи слева). По данному соотношению и определяется сдвиг лейкоцитарной формулы влево и вправо соответственно. Норма нейтрофилов в крови Возраст Норма нейтрофилов (NEUT%) Палочкоядерные Сегментоядерные У взрослых: 1-4 40-60 Нейтрофилы. Большинство лейкоцитов крови (40-60%) является нейтрофилами. Диаметр нейтрофилов - 10-15 мкм. После выхода из костного мозга нейтрофилы в крови циркулируют в среднем около 8 часов. Затем они оставляют русло крови и в течение нескольких суток находятся среди соединительнотканных элементов большинства органов. Здесь они способны захватывать и переваривать (фагоцитировать) микроорганизмы, которые могут попасть в ткани при повреждении. За эту возможность и свои относительно небольшие размеры нейтрофилы называются микрофаги. Пищеварение фагоцитированных микроорганизмов происходит под действием различных факторов, содержащихся в их гранулах. Содержание гранул может инактивировать широкий спектр микрофлоры, вирусов, микоплазм. Особенно активны миелопероксидазы, активирована пероксидом водорода, и лизоцим, гидролизует гликопротеиды бактериальной оболочки. Фагоцитин, катионные белки гранул гидролазы, полипептидазы, ДНКазы и РНКазы вместе с упомянутыми выше соединениями могут расщепить фагоцитированные бактерии любой структуры. Вместе с другими погибшими клетками нейтрофилы образуют основу навоза. Нейтрофилы участвуют также в образовании интерферона - вещества, которое действует на вирусы. В русле крови содержится незначительное количество зрилид клеток. В 20-40 раз больше их е в органах-депо, в частности кровотворчих - костном мозге, селезенке, печени, а также в капиллярах легких. После образования зрелых нейтрофилов еще в течение 5-7 суток остается в костном мозге. Отсюда Нейтрофилы могут выходить и пополнять пул циркулирующих клеток, которые накопились у места повреждения, в области воспаления. Содержание нейтрофилов в крови может увеличиваться как за счет интенсивного лейкопоэза, так и путем перераспределительной реакции, возникающей при эмоциях, физической нагрузке, пищеварении, различных стрессах. Активный выход лейкоцитов из костного мозга приводит к появлению в русле крови юных форм: палочкоядерных или даже метамиелоциты. Рост их количества в крови свидетельствует об активизации процессов создания и выхода нейтрофилов из костного мозга. Перераспределительный лейкоцитоз является следствием не только выхода нейтрофилов из органов-депо, но и изменения средства циркуляции. В обычных условиях в гумне крови нейтрофилы делятся на две части - пул клеток, циркулирующих по центру сосудов, и пул пристеночных клеток. При вышеуказанных состояниях лейкоциты отходят от стенок и присоединяются к циркулирующих клеток. Базофилы составляют 0,5% лейкоцитов крови. Это клетки с сегментированным ядром диаметром 10-12 мкм. Они содержат большое количество таких биологически активных веществ, как гистамин, повышающий проницаемость стенок капилляров, и гепарин, который предотвращает свертыванию крови. В начальный период поступления в организм микроорганизмов базофилы способствуют развитию воспаления, а после ликвидации патологического процесса участвуют в рассасывании очага воспаления. Базофилы являются источником брадикинина, серотонина и ряда лизосомальных ферментов. Они участвуют в аллергических реакциях (измененном иммунитете). Поэтому базофилия является одним из признаков сенсибилизации организма при аллергии. Реснички в реснитчатых клетках эпителия трахеи. Электронные микрофотографии. 1. На снимках приведены срезы ресничек, находящихся на апикальной поверхности реснитчатых клеток эпителия трахеи; причём, сверху - поперечный срез, а снизу - продольный. 2. Ресничка - это покрытое плазмолеммой выпячивание цитоплазмы, по центру которого проходит аксонема (осевая нить). 3. а) Аксонема образована микротрубочками по схеме: (9 ґ 2) + 2 . б) Это значит, что по её окружности расположены девять периферических дуплетов микротрубочек (3), а ещё пара центральных микротрубочек (4) идёт вдоль оси аксонемы. 4. При этом от каждого периферического дуплета на разных его уровнях отходят по направлению к соседнему дуплету - две т.н. ручки из белка динеина, а по направлению к центральному футляру - радиальные мостики. 5. При замыкании и размыкании динеиновых мостиков соседние дуплеты несколько перемещаются друг относительно друга, что приводит к изгибу (биению) реснички. 6. а) Аксонема прикреплена к базальному телу (5), находящемуся в поверхностных слоях цитоплазмы. б) По строению базальное тело похоже на центриоль, т.е. состоит из 9 периферических триплетов. в) При этом по две микротрубочки каждого триплета переходят в дуплет аксонемы. 7. а) Кроме трахеи, реснитчатые клетки имеются во всех других отделах воздухоносных путей дыхательной системы, а также в эпителии половых трактов: - женского (яйцеводы и матка) и - мужского (выносящие канальцы яичка и проток придатка яичка). б) А у сперматозоидов имеются не реснички, а один жгутик; но в основе его - аксонема, организованная так же, как аксонема ресничек. 8. Прочие структуры, видимые на снимках: плазмолемма (1), окружающая ресничку и клетку в целом; цитоплазма реснички (2) и тела клетки (6). Железистые эпителиальные клетки могут располагаться поодиночке, но чаще формируют железы. Клетки железистого эпителия — гландулоциты или железистые клетки, процесс секреции в них протекает циклически, называется секреторным циклом и включает в себя пять стадий: 1. Фаза поглощения исходных веществ (из крови или межклеточной жидкости), из которых образуются конечный продукт (секрет); 2. Фаза синтеза секрета связана с процессами транскрипции и трансляции, деятельностью грЭПС и агрЭПС, комплекса Гольджи. 3. Фаза созревания секрета происходит в аппарате Гольджи: происходит дегидратация и присоединение дополнительных молекул. 4. Фаза накопления синтезируемого продукта в цитоплазме железистых клеток обычно проявляется нарастанием содержания секреторных гранул, которые могут заключаться в мембраны. 5. Фаза выведения секрета может осуществляться несколькими путями: 1) без нарушения целостности клетки (мерокриновый тип секреции), 2) с разрушением апикальной части цитоплазмы (апокриновый тип секреции), с полным нарушением целостности клетки (голокриновый тип секреции). Железы делятся на две группы: 1) железы внутренней секреции, или эндокринные, которые продуцируют гормоны – вещества, обладающие высокой биологической активностью. Выводные протоки отсутствуют, секрет поступает через капилляры в кровь; и 2) железы внешней секреции, или экзокринные, секрет в которых выделяется во внешнюю среду. Экзокринные железы состоят из концевых (секреторных отделов) и выводных протоков.
|
|
|
|
|
|
|
|
|
|
|
|
Строение экзокринных желёз
Концевые (секреторные) отделы состоят из железистых клеток (гландулоцитов), которые продуцируют секрет. Клетки расположены на базальной мембране, для них характерна выраженная полярность: плазмолемма имеет различное строение на апикальных (микроворсинки), базальных (взаимодействие с базальной мембраной) и латеральных (межклеточные контакты) поверхностях клеток. В апикальной части клеток присутствуют секреторные гранулы. В клетках, которые вырабатывают секреты белкового характера (например: пищеварительные ферменты), хорошо развита грЭПС. В клетках, синтезируемых небелковые секреты (липиды, стероиды), выражена аЭПС.
В некоторых железах, образованных эпителиями эпидермального типа (например, потовых, молочных, слюнных), концевые отделы помимо железистых клеток содержат миоэпителиальные клетки – видоизмененные эпителиоциты с развитым сократительным аппаратом. Миоэпителиальные клетки своими отростками охватывают снаружи железистые клетки и, сокращаясь, способствуют выделению секрета из клеток концевого отдела.
Выводные протоки связывают секреторные отделы с покровными эпителиями и обеспечивают выделение синтезированных веществ на поверхность тела или в полость органов.
Разделение на концевые отделы и выводные протоки затруднено в некоторых железах (например, желудка, матки), так как все участки этих простых желез способны к секреции.
Классификация экзокринных желез
I. Морфологическая классификация экзокринных желез основана на структурном анализе их концевых отделов и выводных протоков.
- В зависимости от формы секреторного (концевого) отдела различают альвеолярные, трубчатые и смешанные (альвеолярно-трубчатые) железы;
- В зависимости от ветвления секреторного отдела различают разветвленные и неразветвленные железы.
- Ветвление выводных протоков определяет деление желез на простые (проток не ветвится) и сложные (проток ветвится).
II. По механизму (способу) выведения секрета экзокринные железы делят на апокриновые (молочная железа), голокриновые (сальная железа) и мерокриновые (большинство желез).
Примеры классификации желез. Классификационная характеристикасальной железыкожи: 1) простая альвеолярная железа с разветвленными концевыми отделами, 2) липидная – по химическому составу секрета, 3) голокриновая – по способу выведения секрета.
 Центриоль - внутриклеточный органоид эукариотической клетки, представляющий тельца в структуре клетки, размер которых находится на границе разрешающей способности светового микроскопа.
Центриоль - внутриклеточный органоид эукариотической клетки, представляющий тельца в структуре клетки, размер которых находится на границе разрешающей способности светового микроскопа.
Эти органеллы в делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В неделящихся клетках центриоли часто определяют полярность клеток эпителия и располагаются вблизи комплекса Гольджи. Чаще всего пара центриолей лежит вблизи ядра. Каждая центриоль построена из цилиндрических элементов, образованных в результате полимеризации белка тубулина. Девять триплетов микротрубочек расположены по окружности.
Центриоли принимают участие в формировании цитоплазматических микротрубочек во время деления клетки и в регуляции образования митотического веретена. В клетках высших растений и большинства грибов центриолей нет, и митотическое веретено образуется там иным способом. Кроме того, ученые полагают, что ферменты клеточного центра принимают участие в процессе перемещения дочерних хромосом к разным полюсам в анафазе митоза.
 Лимфоциты составляют 20-40% лейкоцитов. Эти мононуклеары, в отличие от большинства других клеток крови, сохранили способность к пролиферации и дифференцировки.
Лимфоциты составляют 20-40% лейкоцитов. Эти мононуклеары, в отличие от большинства других клеток крови, сохранили способность к пролиферации и дифференцировки.
Возникнув в костном мозге, лимфоциты выходят в кровоток. Но большинство из них имеют повторно созреть в лимфо идних органах. Различают первичные и вторичные лимфо й дни органы. Одним из первичных органов является загрудинная железа (тимус). В период онтогенетического развития она претерпевает существенные изменения. Наибольших размеров достигает в период полового созревания, после чего постепенно инволюционуе. Лимфоциты, как и другие клетки крови, развиваются из общих стволовых клеток-предшественников, часть которых еще в эмбриональный период оставляет костный мозг и оседает в загрудинной железы. Эти клетки дают начало Т-лимфоцитам.
К первичным лимфоидных органов у человека относится костный мозг, а у птиц - Bursa Fabricii. Лимфоциты, которые здесь созревают, относятся к В-лимфоцитов (от лат. Bursa - сумка).
К вторичным органам относятся лимфатические узлы, селезенка и система лимфоэпителиальных образований, которая объединяет скопления лимфоидной ткани в слизистых оболочках желудка и кишечника, дыхательных и мочеполовых путей. В эти образования лимфоциты поступают из костного мозга и тимуса, т.е. это В-и Т-лимфоциты. Во вторичных лимфоидных органах происходит пролиферация лимфоцитов в ответ на поступление в организм чужеродного белка-антигена. Указанные Т-, а возможно, и В-лимфоциты способны к рециркуляции: они снова появляются в крови и поступают в новых участков вторичных лимфоидных органов, где образуют колонии, которые синтезируют пи антитела.
Лимфоциты также участвуют в реакциях клеточного иммунитета-уничтожают клетки, подвергшиеся мутации.

Макрофаги — это вид белых кровяных клеток, которые вместе с промоцитами, монобластами и со своими предшественниками, моноцитами, образуют систему монуклеарных фагоцитов. Фагоциты, в переводе с греческого, означают "есть клетку" (phago - есть, cyt - клетка).
Моноциты и нейтрофилы - два самых основных типа клеток крови, которые способны вбирать в себя и уничтожать чужеродные микроорганизмы. В период, когда моноциты из крови попадают в ткани организма, они превращаются в макрофаги.
По своим функциям моноциты и макрофаги очень близки, так как макрофаги принимают непосредственное участие в воздействии на естественный и приобретенный иммунитеты организма, на секрецию различных биологически активных веществ.
Макрофаги входят в важную группу долгоживущих клеток, способных к фагоцитозу (активному захвату и поглощению живых клеток и отмерших частиц одноклеточными организмами или специальными клетками - фагоцитами).
Воздействие макрофагов на естественный иммунитет заключается в их способности к фагоцитозу и в участии в синтезе активных веществ, таких как пищеварительные ферменты, компонентов систем фагоцитина, интерферона, лизоцима и др.Участие в приобретенном иммунитете проявляется в пассивной передаче антигена иммунокомпетентным клеткам - Т- и В - лимфоцитам, а так же в индукции специфической реакции на антигены.
Кроме кровотока, макрофаги различных типов могут существовать в разных органах: в капиллярах кроветворных органов, в лимфатических узлах, в печени, в легочных альвеолах, в соединительной и в нервной ткани, в костях и в костном мозге.
Основными задачами макрофагов является борьба с бактериями, вирусами, простейшими микроорганизмами и другими внутриклеточными паразитами, и их уничтожение.
Они играют важную роль при сопротивлении раковым заболеваниям, и при значительной стимуляции иммунитета, могут способствовать уничтожению раковых клеток.
При аутоиммунных заболеваниях макрофаги убирают из крови иммунные комплексы.Так же они активно участвуют в заживлении ран и удалении омертвевших клеток.
 Фибробласты— клетки, синтезирующие компоненты межклеточного вещества: белки (например, коллаген, эластин), протеогликаны, гликопротеины. В эмбриональном периоде ряд мезенхимных клеток зародыша дают начало дифферону фибробластов, к которому относят: стволовые клетки, полустволовые клетки-предшественники, малоспециализированные фибробласты, дифференцированные фибробласты (зрелые, активно функционирующие), фиброциты (дефинитивные формы клеток), миофибробласты и фиброкласты.
Фибробласты— клетки, синтезирующие компоненты межклеточного вещества: белки (например, коллаген, эластин), протеогликаны, гликопротеины. В эмбриональном периоде ряд мезенхимных клеток зародыша дают начало дифферону фибробластов, к которому относят: стволовые клетки, полустволовые клетки-предшественники, малоспециализированные фибробласты, дифференцированные фибробласты (зрелые, активно функционирующие), фиброциты (дефинитивные формы клеток), миофибробласты и фиброкласты.
С главной функцией фибробластов связаны образование основного вещества и волокон (что ярко проявляется, например, при заживлении ран, развитии рубцовой ткани, образовании соединительнотканной капсулы вокруг инородного тела).
Фибробласты – это подвижные клетки. В их цитоплазме, особенно в периферическом слое, располагаются микрофиламенты, содержащие белки типа актина и миозина. Движение фибробластов становится возможным только после их связывания с опорными фибриллярными структурами с помощью фибронектина — гликопротеина, синтезируемого фибробластами и другими клетками, обеспечивающего адгезию клеток и неклеточных структур. Во время движения фибробласт уплощается, а его поверхность может увеличиться в 10 раз. По способности синтезировать фибриллярные белки к семейству фибробластов можно отнести ретикулярные клетки ретикулярной соединительной ткани кроветворных органов, а также хондробласты и остеобласты скелетной разновидности соединительной ткани.
 Комплекс Гольджи - это мембранная структура, присущая любой эукариотической клетке.
Комплекс Гольджи - это мембранная структура, присущая любой эукариотической клетке.
Комплекс Гольджи состоит из уплощенных цистерн, как правило, собранных в стопки (диктиосомы). Цистерны не изолированы, а соединены между собой системой трубочек. Первую от ядра цистерну называют цис-полюсом комплекса Гольджи, а последнюю, соответственно, транс-полюсом. Количество цистерн в разных клетках разных организмов может варьировать, но в целом строение комплекса Гольджи у всех эукариот примерно одинаково. В секреторных клетках он развит особенно сильно.
Функции комплекса Гольджи заключаются в переносе белков к месту назначения, а также их гликозилировании, дегликозилировании и модификации олигосахаридных цепочек.
Комплексу Гольджи свойственна функциональная анизотропия. Новосинтезированные белки транспортируются из эндоплазматического ретикулума к цис-полюсу диктиосом с помощью везикул. Далее они постепенно продвигаются по направлению к транс-полюсу, подвергаясь поэтапным модификациям (по мере удаления от ядра состав ферментных систем в цистернах меняется). И, наконец, белки отправляются к своему окончательному месту назначения в везикулах, отпочковывающихся от транс-полюса.
Комплекс Гольджи обеспечивает транспорт белков в три компартмента: к лизосомам (а также центральной вакуоли растительной клетки и сократительным вакуолям простейших), к клеточной мембране и в межклеточное пространство. Направление переноса белка определяется специальными гликозидными метками. Например, маркер лизосомальных ферментов - манноза-6-фосфат. Созревание и транспорт митохондриальных, ядерных и хлоропластных белков происходит без участия комплекса Гольджи: они синтезируются свободными рибосомами после чего попадают непосредственно в цитозоль.
Важная функция комплекса Гольджи - синтез и модификация углеводного компонента гликопротеинов, протеогликанов и гликолипидов. В нем же синтезируются и многие полисахариды, например гемицеллюлоза и пектин у растений. Цистерны комплекса Гольджи содержат целый набор различных гликозилтрансфераз и гликозидаз. Также в них происходит сульфатирование углеводных остатков.
Плазматические клетки встречаются в ликворе только при патологических процессах. Образуются плазматические клетки из В-лимфоцитов в фолликулах корковой зоны лимфатических узлов и краевой зоны белой пульпы селезенки, где при встрече с антигеном они проходят этап антигензависимой дифференцировки. Дифференцировка В-лимфоцитов в плазмобласты продолжается 6-12 ч. Затем после нескольких делений плазмобласт превращается в проплазмоцит, из которого и образуется зрелая плазматическая клетка. Основная функция плазматических клеток - синтез и секреция антител, во время этих процессов в клетках повышается синтез белков, что сказывается на морфологии их цитоплазмы, которая становится похожа на цитоплазму секретирующих клеток.
Плазматические клетки при окраске реактивом Самсона имеют правильную круглую форму, диаметр до 12 мкм. Ядро также правильной круглой формы, окрашивается в темно-вишневый цвет. При увеличениях х900 и х1000 (иммерсия) хорошо видна мелкоглыбчатая или колесовидная структура ядра. Ядро располагается эксцентрично. Цитоплазма обильная, окружает ядро больше с одной стороны, окрашивается реактивом Самсона в ярко-розовый цвет (рис. 141, А, Б, В).
В ликворе иногда встречаются 2-ядерные плазматические клетки. При окраске азур-эозином плазматические клетки в ликворе идентичны плазматическим клеткам разной степени зрелости в периферической крови и костном мозге (рис. 142, А, Б, В).
 Молодая плазматическая клетка (плазмобласт, проплазмоцит)имеет диаметр 16-20 мкл. Ядро занимает большую часть клетки, расположено центрально или несколько эксцентрично, хроматиновой структуры, темно-фиолетового цвета. Более молодые клетки (плазмобласты) содержат 1-2 нуклеолы. Цитоплазма темно-базо-фильная с грязноватым оттенком и с зоной просветления вокруг ядра. Иногда в ликворе зона просветления вокруг ядра у плазматических клеток отсутствует. Клетка, образующаяся при дифференцировке В-лимфоцитов и превращающаяся в проплазмоцит; характеризуется крупным ядром, интенсивно базофильной цитоплазмой с перинуклеарной зоной просветления.
Молодая плазматическая клетка (плазмобласт, проплазмоцит)имеет диаметр 16-20 мкл. Ядро занимает большую часть клетки, расположено центрально или несколько эксцентрично, хроматиновой структуры, темно-фиолетового цвета. Более молодые клетки (плазмобласты) содержат 1-2 нуклеолы. Цитоплазма темно-базо-фильная с грязноватым оттенком и с зоной просветления вокруг ядра. Иногда в ликворе зона просветления вокруг ядра у плазматических клеток отсутствует. Клетка, образующаяся при дифференцировке В-лимфоцитов и превращающаяся в проплазмоцит; характеризуется крупным ядром, интенсивно базофильной цитоплазмой с перинуклеарной зоной просветления.
 Зрелая плазматическая клетка (плазмоцит) Плазматические клетки встречаются в ликворе только при патологических процессах. Образуются плазматические клетки из В-лимфоцитов в фолликулах корковой зоны лимфатических узлов и краевой зоны белой пульпы селезенки, где при встрече с антигеном они проходят этап антигензависимой дифференцировки. Дифференцировка В-лимфоцитов в плазмобласты продолжается 6-12 ч. Затем после нескольких делений плазмобласт превращается в проплазмоцит, из которого и образуется зрелая плазматическая клетка. Основная функция плазматических клеток - синтез и секреция антител, во время этих процессов в клетках повышается синтез белков, что сказывается на морфологии их цитоплазмы, которая становится похожа на цитоплазму секретирующих клеток.
Зрелая плазматическая клетка (плазмоцит) Плазматические клетки встречаются в ликворе только при патологических процессах. Образуются плазматические клетки из В-лимфоцитов в фолликулах корковой зоны лимфатических узлов и краевой зоны белой пульпы селезенки, где при встрече с антигеном они проходят этап антигензависимой дифференцировки. Дифференцировка В-лимфоцитов в плазмобласты продолжается 6-12 ч. Затем после нескольких делений плазмобласт превращается в проплазмоцит, из которого и образуется зрелая плазматическая клетка. Основная функция плазматических клеток - синтез и секреция антител, во время этих процессов в клетках повышается синтез белков, что сказывается на морфологии их цитоплазмы, которая становится похожа на цитоплазму секретирующих клеток.
Плазматические клетки при окраске реактивом Самсона имеют правильную круглую форму, диаметр до 12 мкм. Ядро также правильной круглой формы, окрашивается в темно-вишневый цвет. При увеличениях х900 и х1000 (иммерсия) хорошо видна мелкоглыбчатая или колесовидная структура ядра. Ядро располагается эксцентрично. Цитоплазма обильная, окружает ядро больше с одной стороны, окрашивается реактивом Самсона в ярко-розовый цвет (рис. 141, А, Б, В).
В ликворе иногда встречаются 2-ядерные плазматические клетки. При окраске азур-эозином плазматические клетки в ликворе идентичны плазматическим клеткам разной степени зрелости в периферической крови и костном мозге (рис. 142, А, Б, В).
Молодая плазматическая клетка (плазмобласт, проплазмоцит) имеет диаметр 16-20 мкл. Ядро занимает большую часть клетки, расположено центрально или несколько эксцентрично, хроматиновой структуры, темно-фиолетового цвета. Более молодые клетки (плазмобласты) содержат 1-2 нуклеолы. Цитоплазма темно-базо-фильная с грязноватым оттенком и с зоной просветления вокруг ядра. Иногда в ликворе зона просветления вокруг ядра у плазматических клеток отсутствует.
Зрелая плазматическая клетка (плазмоцит) имеет диаметр 10-12 мкл, круглую форму. Ядро занимает меньшую часть клетки, располагается эксцентрично, структура ядра зрелая с четким делением на окси- и базихроматин. Базихроматин располагается в ядре в виде мелких глыбок (булыжная мостовая) или колесовидно. Базофилия цитоплазмы с сероватым оттенком обычно более выражена по периферии. Иногда просматривается зона просветления цитоплазмы вокруг ядра. Плазматические клетки в ликворе могут быть овальной и неправильной формы с большой, маленькой или отсутствующей зоной просветления вокруг ядра. Иногда в цитоплазме по ее периферии видны единичные вакуоли как признак секреции.

Миофибриллы (с лат.- волокно,ниточка) - это сократимые волокна, находящиеся в структуре сердца и поперечно-полосатых мышц.
Состоят миофибриллы из протофибрилл. Сократимые единицы миофибрилл, хорошо различимы в световом микроскопе. Миофибриллы выражены поперечной исчерченностью, в них можно найти очерёдные области с разным коэффициентом светопреломления (темные анизотропные диски А чередуются со светлыми изотропными дисками I), что придает поперечно-полосатым мышцам свойственный для них вид. Так же делятся миофибриллы на саркомеры,что придают мышцам полосатость. По сути, это сократительные мышцы.
Диаметр миофибриллы достигает нескольких мкм. Основная часть миофибриллы представляят собой тончайшие нити - миофиламенты. Они разделяются на два типа: актиновые (состоящие из актина, длина этих нитей достигает от 1000, до 1100 нанометров) и миозиновые (состоят из миозина, длина 1500 нанометров). На продольном разрезе мышцы, видны чередующие светлые и тёмные полосы. Тёмные - это А-диски, светлые- I-диски.

Тучная клетка – это клетка в тканях тела, содержащая группы биологически активных веществ, которые, высвобождаясь, вызывают симптомы аллергии. Тучные клетки сосредоточены в выстилке носа, пазухах, нижних дыхательных путях и в коже. На поверхности тучных клеток имеется множество рецепторных участков, где присутствует антитело IgE. Подсчитано, что на каждой тучной клетке находится около 300 тысяч таких участков.
Каждая тучная клетка содержит мельчайшие «пакеты» гранул с биологически активными веществами, и каждая окружена собственной мембраной. Когда аллерген объединяется с IgE-антителом, фиксированным на тучной клетке, происходят изменения в клеточной мембране. Это, в свою очередь, вызывает выброс из гранул биологически активных веществ, называемых медиаторами, процесс носит называние дегрануляции.
Именно эти вещества, выделившиеся из тучных клеток, взывают симптомы аллергических реакций, особенно воспаление. Одним из первых идентифицированных медиаторов был гистамин. Природа других, более мощных, длительно действующих веществ, прояснилась только в 1980-х годах. Было обнаружено, что клеточные мембраны тучных клеток и других тканевых клеток, находящихся рядом, выделяют арахидоновые кислоты, которые, в свою очередь, являются исходным материалом для синтеза более мощных соединений, таких как лейкотриены и простагландины.
Известно, что лейкотриены в тысячи раз сильнее гистамина и обладают более продолжительным эффектом. Простагландинов насчитывается двадцать тысяч различных видов, и они производят в организме самые разные эффекты. Эти соединения участвуют в регулировании иммунного ответа, особенно при воспалении. Простагландины, образующиеся вследствие реакции тучной клетки, ответственны за поддержание атак на «захватчика».
Комбинация веществ, выделяемых тучными клетками, играет роль в возникновении трёх реакций, проявляющихся при аллергическом ответе. К ним относятся:
- расширение мелких кровеносных сосудов с повышением проницаемости (или «протекаемости») – основа для аллергической сыпи, отёка Квинке (гигантской крапивницы), заложенности носа и аллергической головной боли;
- спазм гладкой мышцы, вызывающий сжатие дыхательных путей, что типично для астмы; кроме того, он, вероятно, является причиной спазмов, сопровождающих желудочно-кишечную аллергию;
- обильные выделения, очевидные при аллергическом конъюнктивите, слуховых нарушениях, астме и сенной лихорадке.
 Эритроциты – форменные элементы крови, содержащие в себе гемоглобин. Основная функция эритроцитов – перенос кислорода. Нормальный эритроцит имеет двояковогнутую форму. Благодаря такой форме увеличивается площадь поверхности эритроцита, облегчается связывание эритроцита с кислородом. Средний цикл жизни эритроцита 120 дней. Нормы количества RBC (норма эритроцитов крови): Мужчины: 4.5-5.5*1012/л Женщины: 4.0-5.0*1012/л
Эритроциты – форменные элементы крови, содержащие в себе гемоглобин. Основная функция эритроцитов – перенос кислорода. Нормальный эритроцит имеет двояковогнутую форму. Благодаря такой форме увеличивается площадь поверхности эритроцита, облегчается связывание эритроцита с кислородом. Средний цикл жизни эритроцита 120 дней. Нормы количества RBC (норма эритроцитов крови): Мужчины: 4.5-5.5*1012/л Женщины: 4.0-5.0*1012/л
Повышение количества эритроцитов в крови называется эритроцитозом. Эритроцитозы бывают абсолютными и относительными. Абсолютные эритроцитозы возникают при увеличении числа эритроцитов. Относительный эритроцитоз возникает при сгущении крови (уменьшении ее объема).
 Пониженное количество эритроцитов называется эритропенией. Эритропения возникает, к примеру, при кровотечении.
Пониженное количество эритроцитов называется эритропенией. Эритропения возникает, к примеру, при кровотечении.
Полирибосома — несколько рибосом, одновременно транслирующих одну молекулу иРНК. Поскольку длина средней молекулы мРНК значительно превышает количество нуклеотидов, занимаемых на РНК рибосомой, одну молекулу РНК, в зависимости от скорости инициации одновременно транслируют несколько рибосом. Образование и количество рибосом в полисоме зависит от скорости инициации, элонгации и терминации на данной конкретной РНК.
 Как показано при использовании электронной микроскопии, ядро окружено двумя параллельными мембранами, между которыми находится узкое (40—70 нм) пространство — перинуклеарная цистерна. В совокупности парные мембраны и разделяющее их пространство образуют ядерную оболочку. С внутренней мембраной ядерной оболочки тесно связана белковая структура — ядерная фиброзная пластинка, или ядерная ламина, которая придает прочность ядерной оболочке. Фиброзная пластинка образована тремя главными белками, которые называются ламины А, В и С. В неделящихся клетках с ядерной фиброзной пластинкой связаны хромосомы. Характер этой связи в пределах каждой ткани остается неизменным от клетки к клетке, что подтверждает мнение о фиксированном расположении хромосом внутри ядра. К наружной мембране прикреплены полирибосомы, это показывает, что ядерная оболочка является частью ЭПС. Белки, синтезированные на полирибосомах, прикрепленных к ядерной оболочке, временно сегрегируются в перинуклеарной цистерне. В тех участках, где внутренняя и наружная мембраны ядерной оболочки сливаются друг с другом, имеются отверстия — ядерные поры, которые образуют контролируемые пути, связывающие ядро и цитоплазму. Поры являются не простыми отверстиями, а содержат восьмиугольный поровый комплекс, процесса, опосредованного рецепторами, который использует энергию АТФ и осуществляется в два этапа. На первом этапе белки, содержащие один или несколько сигналов ядерной локализации, связываются со специфическими цитозольными белками, образуя комплекс, который временно прикрепляется к комплексу ядерной поры без затраты энергии. На втором этапе белки с сигналом ядерной локализации переносятся в ядро при использовании энергии АТФ, а цитозольный белок остается в цитоплазме. По крайней мере часть энергии АТФ затрачивается на то, чтобы открыть комплекс ядерный поры, что необходимо для осуществления транспорта крупной молекулы. Значительно меньше известно о переносе из ядра в цитоплазму крупных молекул и молекулярных комплексов, например субъединиц рибосом.
Как показано при использовании электронной микроскопии, ядро окружено двумя параллельными мембранами, между которыми находится узкое (40—70 нм) пространство — перинуклеарная цистерна. В совокупности парные мембраны и разделяющее их пространство образуют ядерную оболочку. С внутренней мембраной ядерной оболочки тесно связана белковая структура — ядерная фиброзная пластинка, или ядерная ламина, которая придает прочность ядерной оболочке. Фиброзная пластинка образована тремя главными белками, которые называются ламины А, В и С. В неделящихся клетках с ядерной фиброзной пластинкой связаны хромосомы. Характер этой связи в пределах каждой ткани остается неизменным от клетки к клетке, что подтверждает мнение о фиксированном расположении хромосом внутри ядра. К наружной мембране прикреплены полирибосомы, это показывает, что ядерная оболочка является частью ЭПС. Белки, синтезированные на полирибосомах, прикрепленных к ядерной оболочке, временно сегрегируются в перинуклеарной цистерне. В тех участках, где внутренняя и наружная мембраны ядерной оболочки сливаются друг с другом, имеются отверстия — ядерные поры, которые образуют контролируемые пути, связывающие ядро и цитоплазму. Поры являются не простыми отверстиями, а содержат восьмиугольный поровый комплекс, процесса, опосредованного рецепторами, который использует энергию АТФ и осуществляется в два этапа. На первом этапе белки, содержащие один или несколько сигналов ядерной локализации, связываются со специфическими цитозольными белками, образуя комплекс, который временно прикрепляется к комплексу ядерной поры без затраты энергии. На втором этапе белки с сигналом ядерной локализации переносятся в ядро при использовании энергии АТФ, а цитозольный белок остается в цитоплазме. По крайней мере часть энергии АТФ затрачивается на то, чтобы открыть комплекс ядерный поры, что необходимо для осуществления транспорта крупной молекулы. Значительно меньше известно о переносе из ядра в цитоплазму крупных молекул и молекулярных комплексов, например субъединиц рибосом.
 Ядрышко находится внутри ядра, и не имеет собственной мембранной оболочки, однако хорошо различимо под световым и электронным микроскопом.
Ядрышко находится внутри ядра, и не имеет собственной мембранной оболочки, однако хорошо различимо под световым и электронным микроскопом.
Основной функцией ядрышка является синтез рибосом. В геноме клетки имеются специальные участки, так называемые ядрышковые организаторы, содержащие гены рибосомной РНК (рРНК) , вокруг которых и формируются ядрышки. В ядрышке происходит синтез рРНК РНК полимеразой I, ее созревание, сборка рибосомных субчастиц. В ядрышке локализуются белки, принимающие участие в этих процессах. Некоторые из этих белков имеют специальную последовательность — сигнал ядрышковой локализации (NoLS, от англ. Nucleolus Localization Signal). Следует отметить, самая высокая концентрация белка в клетке наблюдается именно в ядрышке. В этих структурах было локализовано около 600 видов различных белков, причем считается, что лишь небольшая их часть действительно необходима для осуществления ядрышковых функций, а остальные попадают туда неспецифически.
Под электронным микроскопом в ядрышке выделяют несколько субкомпартментов. Так называемые Фибриллярные центры окружены участками плотного фибриллярного компонента, где и происходит синтез рРНК. Снаружи от плотного фибриллярного компонента расположен гранулярный компонент, представляющий собой скопление созревающих рибосомных субчастиц.
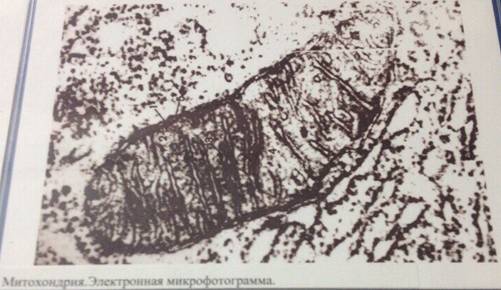 Митохондрии- это органеллы размером с бактерию (около 1 х 2 мкм). Они найдены в большом количестве почти во всех эукариотических клетках. Обычно в клетке содержится около 2000 митохондрий, общий объем которых составляет до 25% от общего объема клетки. Митохондрия ограничена двумя мембранами - гладкой внешней искладчатой внутренней, имеющей очень большую поверхность. Складки внутренней мембраны глубоко входят в матрикс митохондрий, образуя поперечный перегородки - кристы. Пространство между внешней и внутренней мембранами обычно называют межмембранным пространством.
Митохондрии- это органеллы размером с бактерию (около 1 х 2 мкм). Они найдены в большом количестве почти во всех эукариотических клетках. Обычно в клетке содержится около 2000 митохондрий, общий объем которых составляет до 25% от общего объема клетки. Митохондрия ограничена двумя мембранами - гладкой внешней искладчатой внутренней, имеющей очень большую поверхность. Складки внутренней мембраны глубоко входят в матрикс митохондрий, образуя поперечный перегородки - кристы. Пространство между внешней и внутренней мембранами обычно называют межмембранным пространством.
Различный типы клеток отличаются друг от друга как по количеству и форме митохондрий, так и по количеству крист. Особенно много крист имеют митохондрии в тканях с активными окислительными процессами, например в сердечной мышце. Вариации митохондрий по форме, что зависит от их функционального состояния, могут наблюдаться и в тканях одного типа. Митохондрии — изменчивые и пластичные органеллы.
Мембраны митохондрий содержат интегральные мембранные белки. Во внешнюю мембрану входят порины, которые образуют поры и делают мембраны проницаемыми для веществ с молекулярной массой до 10 кДа (см. рис. 223). Внутренняя же мембрана митохондрий непроницаема для большинства молекул; исключение составляют О2, СО2, Н20. Внутренняя мембрана митохондрий характеризуется необычно высоким содержанием белков (75%). В их число входят транспортные белки-переносчики (см. рис. 215), ферменты, компоненты дыхательной цепи иАТФ-синтаза. Кроме того, в ней содержится необычный фосфолипид кардиолипин (см. с. 56). Матрикс также обогащен белками, особенно ферментами цитратного цикла.
Б. Метаболические функции
Митохондрии являются «силовой станцией» клетки, поскольку за счет окислительной деградации питательных веществ в них синтезируется большая часть необходимого клетке АТФ (АТР). В митохондриях локализованы следующие метаболические процессы: превращение пирувата в ацетил-КоА, катализируемое пируватдегидрогеназным комплексом: цитратный цикл; дыхательная цепь, сопряженная с синтезом АТФ (сочетание этих процессов носит название «окислительное фосфорилирование»); расщепление жирных кислот путем β-окисления и частично цикл мочевины. Митохондрии также поставляют клетке продукты промежуточного метаболизма и действуют наряду с ЭР как депо ионов кальция, которое с помощью ионных насосов поддерживает концентрацию Са2+в цитоплазме на постоянном низком уровне (ниже 1 мкмоль/л).
Главной функцией митохондрий является захват богатых энергией субстратов (жирные кислоты, пируват, углеродный скелет аминокислот) из цитоплазмы и их окислительное расщепление с образованием СО2 и Н2О, сопряженное с синтезом АТФ.
Реакции цитратного цикла приводят к полному окислению углеродсодержащих соединений (СО2) и образованию восстановительных эквивалентов, главным образом в виде восстановленных коферментов. Большинство этих процессов протекают в матриксе. Ферменты дыхательной цепи, которые реокисляют восстановленные коферменты, локализованы во внутренней мембране митохондрий. В качестве доноров электронов для восстановления кислорода и образования воды используются НАДН и связанный с ферментом ФАДН2. Эта высоко экзергоническая реакция является многоступенчатой и сопряжена с переносом протонов (Н+) через внутреннюю мембрану из матрикса в межмембранное пространство. В результате на внутренней мембране создаетсяэлектрохимический градиент. В митохондриях электрохимический градиент используется для синтеза АТФ из АДФ (ADP) и неорганического фосфата (Рi) при катализе АТФ-синтазой.
 Внутренняя мембрана митохондрий образует выросты в полость в виде пластин или трубок, называемых кристами. Кристы бывают различных типов. Пространство между кристами заполнено однородным прозрачным веществом - матриксом митохондрий.
Внутренняя мембрана митохондрий образует выросты в полость в виде пластин или трубок, называемых кристами. Кристы бывают различных типов. Пространство между кристами заполнено однородным прозрачным веществом - матриксом митохондрий.
Способность образовывать многочисленные выпячивания является характерной чертой митохондрий. Такие выпячивания ( кристы ) чаще всего имеют вид плоских гребней. Кристы участвуют в преобразовании энергии питательных вещ-в, поступающих в клетку, в энергию молекул.

Эндоплазматическая сеть — это обязательная органелла эукариотической клетки. Она обнаружена в клетках растений, животных и человека. Функции этой составляющей части клетки разнообразны и связаны в основном с синтезом, модификацией и транспортом органических соединений. Впервые эндоплазматическая сеть была обнаружена в 1945 году. Американский ученый К. Гранулярная эндоплазматическая сеть: строение. Данная органелла представляет собой системы цистерн, пузырьков и канальцев. Стенки ее состоят из билипидной мембраны. У мало специализированных клеток, которые характеризируются низким уровнем метаболизма, ЭПС представлена всего лишь несколькими разрозненными цистернами. Внутри клетки, которая активно синтезирует белок, эндоплазматическая сеть состоит из множества цистерн и разветвленной системы канальцев. Как правило, гранулярная ЭПС посредством канальцев связана с мембранами ядерной оболочки — именно таким образом происходят сложные процессы синтеза и транспорта белковых молекул. Гранулярная эндоплазматическая сеть: функции. Как уже упоминалось, вся поверхность ЭПС со стороны цитоплазмы покрыта рибосомами, которые, как известно, участвуют в синтезе белка. ЭПС — это место синтеза и транспорта протеиновых соединений. Что может случится с мозгом из-за мобильного телефона Что в течение часа делает с организмом баночка "Кока-Колы"? О чем больше всего сожалеют люди в конце жизни Эта органелла отвечает за синтез интегральных белков цитоплазматической мембраны. Но в большинстве случаев созданные белковые молекулы далее с помощью мембранных пузырьков транспортируются в комплекс Гольджи, где происходит их дальнейшая модификация и распределение соответственно потребностям клетки и тканей. Кроме того, в полостях цистерн ЭПС происходят и некоторые изменения белка — например, присоединение к нему углеводного компонента. Здесь же, путем агрегации образуются большие секреторные гранулы.
Плазмолемма (цитолемма, плазматическая мембрана) создает селективный барьер между клеткой и внешней средой. Уже в начале XIX в. было обнаружено набухание и сжатие клеток, погруженных в растворы различной ионной силы, что свидетельствовало о наличии на поверхности клетки полупроницаемой мембраны. В 30-е гг. XX в. было доказано, что она состоит из липидов и белков, причем липиды образуют в ней бимолекулярный слой. Плазмолемма непроницаема для макромолекул, поэтому белки цитоплазмы создают в клетке осмотическое давление, под действием которого вода непрерывно поступала бы в клетку, если бы вне клетки не поддерживалась уравновешивающая концентрация других веществ. Это равновесие создается, прежде всего, молекулярным насосом, который выкачивает из клетки ионы натрия и закачивает в клетку ионы калия. З счет разности концентраций ионов внутри и вне клетки плазмолемма приобретает потенциал до +85 мВ.
В электронном микроскопе плазмолемма выглядит как типичная биологическая мембрана, состоящая из двух электронноплотных слоев, между которыми находится электроннопрозрачный слой. Общая толщина всех трех слоев в плазмолемме составляет 12-14 нм. Однако надо отметить, что эта трехслойная структура является лишь основой плазмолеммы, поскольку к ней снаружи и изнутри примыкают слабоконтрастируемые молекулярные комплексы.
Плазмолемма обладает асимметрией, которая выражается в различиях состава липидов и белков наружной и внутренней частей. Асимметрия плазмолеммы проявляется у животных клеток, в частности, в том, что они имеют на своей внешней поверхности особый углеводный слой – гликокаликс. Он образован входящими в состав мембраны олигосахаридами и липидами в комплексе с белками. Гликокаликс присутствует у всех животных клеток, однако степень его развития может быть разной. В наибольшей степени он развит у всасывающих клеток кишечного эпителия. Гликокаликс этих клеток создает среду для пристеночного пищеварения, а также защищает плазмолемму от повреждений. Вирус гриппа содержит фермент нейраминидазу, который необходим для удаления одного из компонентов гликокаликса - сиаловой кислоты, препятствующей прикреплению вирусных частиц к плазмолемме эпителиальных клеток. Гликокаликс обладает выраженными антигенными свойствами, что облегчает опознание клеток при их взаимодействии между собой.
С внутренней стороны плазмолемма связана с такими компонентами цитоскелета, как микротрубочки и микрофиламенты. Это позволяет животной клетке не только поддерживать определенную форму, но и активно изменять ее. Взаимодействие плазмолеммы с цитоскелетом лежит в основе активного движения фибробластов и макрофагов, удлинения проходящих по капиллярам эритроцитов, изменения формы клетки при фагоцитозе и секреции.
Плазмолемма, отграничивая содержимое клетки от внешней среды, одновременно обеспечивает избирательный обмен веществ между клеткой и средой. Транспорт веществ через плазмолемму осуществляется при помощи различных механизмов.
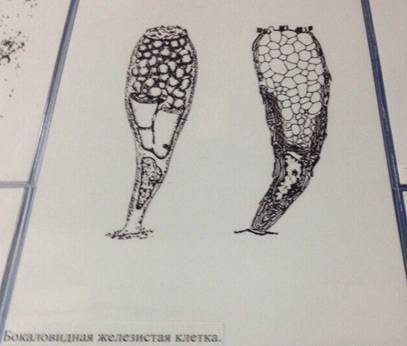 Бокаловидные клетки (синонимы: энтероциты бокаловидные, бокаловидные экзокриноциты; лат. enterocytus caliciformis) — продуцирующие слизь клетки эпителия слизистой оболочки кишечника и других органов позвоночных животных и человека[1]. Бокаловидные клетки часто называют одноклеточными железами[2].
Бокаловидные клетки (синонимы: энтероциты бокаловидные, бокаловидные экзокриноциты; лат. enterocytus caliciformis) — продуцирующие слизь клетки эпителия слизистой оболочки кишечника и других органов позвоночных животных и человека[1]. Бокаловидные клетки часто называют одноклеточными железами[2].
 Локализация
Локализация
Бокаловидные клетки — один из энтероцитов эпителия кишечника. Располагаются на кишечных ворсинках поодиночке среди каёмчатых энтероцитов. Обычно отсутствуют на самой вершине ворсинки, а также на дне крипты. В тонкой кишке человека составляют 9,5 % от всех клеток эпителиального пласта. Число бокаловидных клеток увеличивается в дистальном направлении кишки.[3]
Кроме кишечника, бокаловидные клетки имеются в слизистой оболочке дыхательных путей, в конъюнктиве глаз, протоках поджелудочной и околоушных слюнныхжелёз.[4] В дыхательных путях больше всего бокаловидных клеток находится в экстраторакальной части трахеи, с уменьшением диаметра бронхов их количество существенно сокращается, а вбронхиолах менее 1 мм их нет совсем.[5]
Структура и функции
Бокаловидные клетки накапливают гранулы муциногена, которые, абсорбируя воду, набухают и превращаются в муцин (основной компонент слизи). При этом клетки обретают форму бокала, суженного у основания (где находится ядро) и округлой широкой в апикальной, верхней части. Затем набухшая верхняя часть бокаловидной клетки разрушается, слизь переходит в просвет органа, клетка приобретает призматическую форму и снова начинает накапливать муциноген. Слизь, выделяемая бокаловидными клетками, служит для увлажнения поверхности слизистой оболочки кишечника и этим способствует продвижению химуса, а также участвует в процессах пристеночного пищеварения.
Апикальная часть бокаловидных клеток имеет исчерченную каёмку, подобно каёмчатым энтероцитам. Однако, в отличие от каёмчатых энтероцитов, микроворсинки каёмки у бокаловидных клеток расположены реже и неодинаковы по высоте.
| Волосковая клетка кортиева органа. Схема.
 Мы поможем в написании ваших работ! | |||