Молекулярно-генетические методы
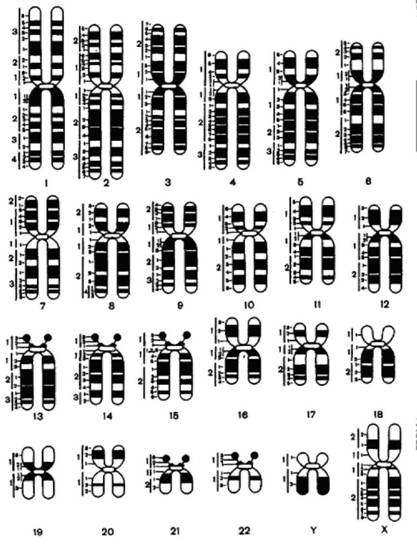
Значительные успехи в области молекулярно-генетического исследования психики человека стали возможны благодаря появлению в 70-х года такого экспериментального инструмента, как рестрикционные эндонуклеазы. Набор специальных ферментов обладает способностью вступать в реакцию с определёнными участками в ДНК, которые называют "сайты узнавания", и разрезать двухцепочечную молекулу ДНК так, что одна из цепей ДНК оказывается на несколько нуклеотидов длиннее другой. Эти нуклеотиды, называемые также "липкими концами", могут спариваться с комплементарными им нуклеотидами. Вследствие этого ДНК разных организмов могут объединяться, образуя так называемые рекомбинантные молекулы. Это свойство используют для размножения специфической, интересующей исследователя ДНК. В 1971 году на Пражской конференции был представлен метод дифференциальной окраски хромосом, благодаря которой каждая хромосома приобретает свой неповторимый рисунок, что помогает точной дифференциации хромосом (рис. 4.6).
Математические методы
К ним относятся методы математической статистики — дисперсионный анализ, анализ корреляций между различными группами людей, анализ достоверности различий между группами.
http://pidruchniki.com/1253121263241/psihologiya/drugie_metody
ГЕНЕТИКА ПОПУЛЯЦИЙ
Понятие популяции в психогенетике
Виды живых существ, населяющие землю, образуют сообщества, то есть пространственно-временные объединения. Одной из разновидностей сообществ является популяция — сообщество одного вида, занимающее определённую территорию. Законы распространения генов среди населения изучает популяционная генетика.
|
|
|
Генетическая характеристика популяций позволяет установить генофонд популяции, факторы и закономерности, обусловливающие сохранение генофонда или его изменение в поколениях. Изучение особенностей распространения психических свойств в разных популяциях даёт возможность прогнозировать распространённость этих свойств в последующих поколениях. Генетическая характеристика популяции начинается с оценки распространённости изучаемого свойства или признака среди населения. По данным о распространённости признака определяются частоты генов и соответствующих генотипов в популяции.
Основными характеристиками генетической популяции являются:
• принадлежность особей одному виду,
• пространственно-временное сходство,
• сходство экологических потребностей,
• способность случайно и свободно скрещиваться между собой — панмиксия. Панмиксия может нарушаться, если образование пар происходит неслучайно. Например, в человеческих популяциях имеется тенденция к неслучайному подбору супружеских пар по росту, интеллекту, интересам и др. Такой неслучайный подбор пар называется ассортативность.
|
|
|
Замкнутая географически или по религиозным соображениям популяция, в которой не происходит обмена особями с другими популяциями, называется изолятом.
Закон Харди-Вайнберга
Зависимости между частотами аллелей и частотами генотипов в поколениях впервые были описаны в 1908 году независимо друг от друга английским математиком Г. Харди и немецким врачом В. Вайнбергом (рис. 5.1). Этот закон определяет взаимоотношения между частотами аллелей в исходной популяции и частотами генотипов в следующем поколении.

Рисунок 5.1. Фотографии и биографические данные Г. Харди и В. Вайнберга
Закон Харди-Вайнберга рассматривает популяцию идеальную. На самом же деле реальная популяция будет не в полной мере соответствовать данному закону, так как в ней происходят такие процессы, влияющие на изменение частот аллелей в популяции, как мутации, миграции, дрейф генов, отбор, а в человеческих популяциях и ассортативность.
Рассмотрим отдельно указанные факторы.
Мутации и виды мутаций
Мутации— внезапные и устойчивые изменения генотипа. Термин "мутация" предложил в 1901 году голландец Гуго де Фриз. Мутации служат основным источником генетической изменчивости, но их частота мала. Для того чтобы мутации привели к значительному изменению частот аллелей, потребуется очень много времени.
|
|
|
Можно классифицировать мутации по различным основаниям. Так, мутации бывают:
• спонтанные и индуцированные, т.е. возникшие под влиянием мутагенов — а) физических излучений; б) химических веществ; в) биологических — влияние вирусов, например, вируса краснухи;
• генные, цитоплазмические, хромосомные и геномные (изменения числа хромосом);
• в зависимости от влияния на жизнеспособность — отрицательные, нейтральные и положительные (роль мутации выявлена в устойчивости к таким заболеваниям, как ВИЧ и серповидноклетчатая анемия);
• в зависимости от типа наследования — доминантные и рецессивные;
• соматические или репродуктивные (гаметные).
Гаметные мутации — мутации, происходящие в половых клетках, например, рак груди. По прогнозу у женщин, родившихся после 1980 года, риск заболеть до 80 лет составляет 12%, то есть заболеет каждая восьмая. Мутированный ген на 13 и 17 хромосоме обусловливает от 5 до 10% случаев рака груди. Ген передаётся по менделевским законам.
|
|
|
Ген ВЯСЛ1, ответственный за генетические формы рака груди, теперь можно назвать геном Анджелины Джоли, поскольку широкой общественности он стал известен благодаря её недавним действиям и публичным заявлениям. Этот ген и его роль в развитии рака известны с середины 90-х годов прошлого века. Причём А. Джоли далеко не первая, кому пришло в голову провести превентивную мастэктомию. Есть данные, что в Великобритании на протяжении 2010-2011 гг. провели около 1500 таких операций именно с превентивной целью.
Надо подчеркнуть, что чисто генетический рак, то есть такой, который возник только из-за унаследованного конкретного "плохого" гена, встречается редко. Как уже говорилось, не больше 10% случаев рака груди и яичников являются наследственными, и за 50% из них отвечают гены ВЯСЛ. Частота мутантного аллеля гена ВЯСЛ1 составляет 0,06%, среди евреев ашкенази больше — 2,6%. Разработано несколько тестов, которые с помощью специальной компьютерной программы рассчитывают риск рака на основании анализа генов ВЯСЛ и индивидуальной информации. Для А. Джоли программа рассчитала наивысший риск рака груди — 86%.
Соматические — остальные 80% мутаций, связанных с возникновением рака груди, что происходят в соматических клетках.
Рассмотрим отдельно виды хромосомных и геномныхмутаций (рис. 5.2).

Рисунок 5.2. Хромосомные мутации
К хромосомным мутациямотносятся делении, дупликации, инверсии, транслокации:
• деления — потеря участка хромосомы;
• дупликация — удвоение;
• транслокация — перенос участка хромосомы на другую;
• инверсия — поворот на 180 градусов определённого участка хромосомы.
Геномные мутациихарактеризуются изменением числа хромосом. Геномные мутации описывают несколькими видами. У человека известны полиплоидия (в том числе тетраплоидия и триплоидия) и анеуплоидия (рис. 5.3).

Рисунок 5.3. Виды геномных мутаций
Полиплоидия— увеличение числа наборов хромосом, кратное гаплоидному (3п, 4п, 5п и т.д.). То есть число хромосом становится равным 69, 92 и т.д. Причины полиплоидии — двойное оплодотворение и отсутствие первого мейотического деления. У человека полиплоидия, а также большинство анеуплоидий приводят к формированию летальных исходов сразу после рождения или до рождения (спонтанные выкидыши).
Анеуплоидия— изменение (уменьшение — моносомияили увеличение — трисомия)числа хромосом в диплоидном наборе, то есть число хромосом, не кратное гаплоидному (2п+1, 2п-1 и т.д.). Число хромосом становится равным 45, 47, 48 и др. Механизмы возникновения анеуплоидий различны: нерасхождение хромосом (хромосомы отходят к одному полюсу, при этом на каждую гамету с одной лишней хромосомой приходится другая — без одной хромосомы) и "анафазное отставание" (в анафазе одна из передвигаемых хромосом отстаёт от всех других).
Трисомия— наличие трёх гомологичных хромосом в кариотипе (например, по 21-й паре, что приводит к развитию синдрома Дауна; по 18-й паре — синдрома Эдвардса; по 13-й паре — синдрома Патау).
Моносомия— наличие только одной из двух гомологичных хромосом. При моносомии по любой из аутосом нормальное развитие эмбриона невозможно. Единственная совместимая с жизнью моносомия у человека — по Х-хромосоме — приводит к развитию синдрома Шерешевского-Тёрнера (45, Х0).
Одним из факторов возникновения мутаций является инбридинг. Инбридинг— кровнородственные браки, например между двоюродными сибсами. В браках между генетическими родственниками повышается вероятность появления потомства с рецессивными признаками. Генетические последствия таких браков проиллюстрируем на примере ряда наследственных болезней в популяциях Европы и США. Например, среди белого населения США на кровнородственные браки проходится лишь 0,05 % от общего числа браков и в то же время 20% случаев альбинизма.
Однако не во всех популяциях последствия инбридинга отрицательны. У сельского населения Индии, Китая и Японии кровнородственные браки довольно часты, но отрицательные эффекты (число уродств, мертворождений) обнаружены не были. Скорее всего в этих странах, где кровнородственные браки разрешены культурой, в течение многих поколений происходило выщепление рецессивных гомозигот, которые обладали пониженной жизнестойкостью.
Миграция и дрейф генов
Миграциейназывается перемещение индивидов из одной популяции в другую с последующим образованием брачных связей между мигрантами и членами исходной популяции. Миграция ведёт к изменению генетического состава популяции, обусловленного поступлением новых генов. Например, распределение группы крови В в Европе является следствием движения монгол в западном направлении от материнской популяции в период между 6 и 15 веками. Поэтому в Европе частоту аллеля В последовательно снижается начиная от границ с Азией и заканчивая Испанией и Португалией. Обмен генами между популяциями может иметь ощутимые медицинские последствия. Так, до недавнего времени резус-конфликт практически не встречался в Китае, так как все китаянки резус-положительны.
Однако процессы миграции, переезд в Китай американцев, межрасовые браки ввели в китайские популяции резус-отрицательный аллель. И если в первом поколении у потомства американцев-мужчин и китаянок-женщин резус-конфликт не наблюдался, однако в последующих частота его встречаемости повысилась, так как появились резус-отрицательные женщины, которые выходили замуж за резус-положительных мужчин.
Вследствие ограниченного числа индивидов, образующих популяцию, возможны случайные изменения частот генов, которые называются дрейфом генов.В ряду поколений, если не действуют другие факторы, дрейф генов может привести к фиксации одного аллеля и исчезновению другого.
С. Райт экспериментально доказал, что в маленьких популяциях частота мутантного аллеля меняется быстро и случайным образом. Его опыт был прост: в пробирки с кормом он посадил по две самки и по два самца мух дрозофил, гетерозиготных по гену А (их генотип можно записать Аа). В этих искусственно созданных популяциях концентрация нормального (А) и мутационного (а) аллелей составила 50%. Через несколько поколений оказалось, что в некоторых популяциях все особи стали гомозиготными по мутантному аллелю (а), в других популяциях он был вовсе утрачен, и, наконец, часть популяций содержала как нормальный, так и мутантный аллель. Важно подчеркнуть, что, несмотря на снижение жизнеспособности мутантных особей и, следовательно, вопреки естественному отбору, в некоторых популяциях мутантный аллель полностью вытеснил нормальный. Это и есть результат случайного процесса — дрейфа генов.
Естественным отбором называется процесс избирательного воспроизводства потомства генетически разными индивидами в популяции. Естественный отбор проявляется в том, что особи с разными генотипами оставляют неодинаковое количество потомства, то есть вносят неодинаковый генетический вклад в следующее поколение.
Таким образом, закон Харди-Вайнберга — это закон популяционной генетики, гласящий, что в популяции бесконечно большого размера, в которой не действует отбор, не идёт мутационный процесс, отсутствует обмен особями с другими популяциями, не происходит дрейф генов, все скрещивания случайны, — частоты генотипов по какому-либо гену (в случае, если в популяции есть два аллеля этого гена) будут поддерживаться постоянными из поколения в поколение и соответствовать уравнению:
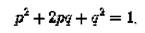
где Р — доля гомозигот по одному из аллелей; Р — частота этого аллеля;
¥^ — доля гомозигот по альтернативному аллелю; Я — частота соответствующего аллеля; — доля гетерозигот.
http://pidruchniki.com/1821011663242/psihologiya/genetika_populyatsiy
НАСЛЕДСТВЕННОСТЬ И ПАТОЛОГИЯ
Прежде всего, необходимо отметить, что термин "наследственные болезни" не тождественен термину "врождённые болезни". Так, врождённые болезни могут быть сформированы под влиянием различных негативных факторов среды, в том числе и внутриутробной.
Всю наследственную патологию можно разбить на 5 групп:
1) генные патологии;
2) хромосомные патологии;
3) болезни с наследственной предрасположенностью;
4) генетические болезни соматических клеток;
5) болезни несовместимости матери и плода.
Генные болезни могут быть моногенными (например, фенилкетонурия) и полигенными (шизофрения), наследоваться по рецессивному или доминантному принципу, быть обусловлены мутацией ядерной или цитоплазмической ДНК.
Дата добавления: 2018-05-02; просмотров: 658; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!