Рис2. Графическая зависимость изменения численности популяций. Фазовый портрет системы
Математическое моделирование.
Моделирование – это исследование явлений и процессов на моделях. Построение модели связано с упрощением изучаемого процесса, в то же время модель должна отражать основные свойства оригинала.
В биологии и медицине используют четыре типа моделей:
1).Биологические. На них изучают общие биологические закономерности, патологические процессы, методы лечения и т.д. (Эксперименты на лабораторных животных).
2).Физические, которые ведут себя аналогично оригиналу. (Аппараты искусственного дыхания, кровообращения и т.д.).
3).Кибернетические – это различные электронные устройства, с помощью которых моделируются информационные процессы в живом организме.( Управление движением руки, ноги, искусственный интеллект и т.д.).
4).Математические. Моделируется математическая зависимость, которая описывает данный процесс. Позволяет
- уменьшить время исследования и число экспериментов;
- давать прогноз течения болезни и подобрать оптимальные варианты лечения;
- судить о таких системах и в таких условиях, которые сложно создать в эксперименте или клинике.
При построенииматематической модели различают 3 этапа:
1). Изучение процесса или явления, сбор данных о его параметрах и создание описательной теории, в которой выявляются причинно-следственные связи, существенные и несущественные детали процесса.
|
|
|
2). Составление математических (чаще всего дифференциальныхуравнений).
3). Рассмотрение на модели возможных результатов и выявление наиболее вероятного исхода, т.е. анализ решения уравнений.
При моделировании сложного процесса приходится составлять систему из большого числа дифференциальных уравнений, которые содержат большое число параметров. Для решения уравнений и определения параметров используют ЭВМ.
Основные направления математического моделирования:
1. Модели проницаемости клеточных мембран (нервная проводимость).
2. Иммунология и эндокринология (ход болезни и лечебные мероприятия).
3. Эпидемиология – распространение инфекционных болезней.
4. Процессы транспорта электронов.
5. Моделирование процесса старения клеток, злокачественных образований.
Математическое моделирование биологических процессов началось с создания первых простейших моделей экологической системы.
Попытки математического моделирования динамики, как отдельных биологических популяций, так и сообществ, включающих взаимодействующие популяции различных видов, предпринимались давно. Одна из первых моделей роста изолированной популяции была предложена ещё в 1798 году Томасом Мальтусом:
|
|
|

Данная модель задаётся следующими параметрами:
N – численность популяции;
μ – разность между коэффициентами рождаемости и смертности.
Модель совместного сосуществования двух биологических видов (популяций) типа «хищник-жертва» впервые была получена американским математиком Альфредом Лоткой в 1925 году. В 1926 году (независимо от Лотки) аналогичные (и более сложные модели) были разработаны итальянским математиком Вито Вольтеррой. Его глубокие исследования в области экологических проблем создали основу математической теории биологических сообществ ( математической экологии).
Поэтому эта классическая математическая модель («хищник-жертва») известна как модель « Лотки-Вольтерра».
Модель Лотки-Вольтерра (хищник-жертва).
Допустим, в некотором замкнутом районе живут рыси и зайцы. Рыси питаются только зайцами, а зайцы - растительной пищей, имеющейся в неограниченном количестве. Необходимо найти макроскопические характеристики, описывающие популяции. Такими характеристиками являются число особей в популяциях - число зайцев N1 и число рысей N2.
Запишем дифференциальные уравнения, описывающие процесс изменения числа особей во времени.
|
|
|
При отсутствии рысей, изменение числа зайцев будет:

α - коэффициент, характеризующий размножение зайцев (жертв).
При отсутствии зайцев, изменение числа рысей будет:

β - коэффициент, характеризующий вымирание рысей (хищников).
При совместном существовании зайцев и рысей:

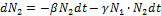
 - коэффициент, характеризующий убыль зайцев, вследствие их встреч с рысями.
- коэффициент, характеризующий убыль зайцев, вследствие их встреч с рысями.
 - коэффициент, характеризующий прирост рысей, вследствие их встреч с зайцами.
- коэффициент, характеризующий прирост рысей, вследствие их встреч с зайцами.
Скорость изменения популяций

 (1)
(1)
Т.е. имеем систему нелинейных дифференциальных уравнений. В стационарном состоянии, когда не изменяется численность зайцев и рысей имеем:
 и
и  и, следовательно,
и, следовательно, 
Т.е. 
Решение этих уравнений (особые точки):

(2)

Отсюда следует:
Вывод1:стационарные состояния не зависят от численности популяции, а определяются только коэффициентами прироста и потерь для другого вида.
Для определения устойчивости в стационарных состояниях необходимо исследовать систему вблизи этих состояний.
Допустим, возникли некоторые случайные отклонения, флуктуации n1, и nг. Определим поведение системы.
|
|
|


Возьмем производные. С учетом того, что производная от стационарного состояния равна 0, получим:

Подставив в (1), раскроем скобки, приведем подобные члены и пренебрежем членами  и
и  вследствие их предполагаемой малости. Результатом пренебрежения ими будет линеаризация уравнений. В результате всех преобразований, окончательно получаем систему линейных дифференциальных уравнений второго порядка типа
вследствие их предполагаемой малости. Результатом пренебрежения ими будет линеаризация уравнений. В результате всех преобразований, окончательно получаем систему линейных дифференциальных уравнений второго порядка типа  , описывающих консервативную колебательную систему, (т.е. идеализированную систему, в которой запас энергии в процессе колебаний остается постоянным):
, описывающих консервативную колебательную систему, (т.е. идеализированную систему, в которой запас энергии в процессе колебаний остается постоянным):

Решив эту систему уравнений получаем 
где: 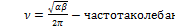

Вывод 2:популяции жертв и хищников испытывают периодические колебания одинаковой частоты , смещенные по фазе (причем максимум численности жертв всегда опережает максимум численности хищников).

Рис 1.Зависимость изменения популяций от времени.
Рассмотрим график зависимости N1от N2, т.е. избавимся от t.
Очевидно, что упрощенное решение нашей системы дифференциальных уравнений путем избавления от элементов и привело нас к тому, что модель пришлось стишком идеализировать, что плохо соответствует реальной модели.
Сделаем попытку решить систему дифференциальных уравнений (1) другим методом. Разделим одно уравнение на другое, тогда получим 
Разделив переменные и проинтегрировав, получим решение:

Константу С можно найти в некоторый момент времени, когда мы знаем N1 и N2 ясно, что она может принимать множество значений.
Итак, мы получили выражение, связывающее две переменные  и
и  , т.е. зависимость =f ( )в неявном виде.
, т.е. зависимость =f ( )в неявном виде.
Начертим график этой функции. Полученная замкнутая кривая не является эллипсом (рис.2 ), хотя отдаленно и напоминает эллипс, который получается при сложении колебаний одинаковой частоты и произвольной фазы.

Рис2. Графическая зависимость изменения численности популяций. Фазовый портрет системы.
Однако и здесь имеют место следующие закономерности:
Вывод 3:
Колебания численности популяций,N1 иN2, действительно имеют место, они сдвинуты по фазе, но не являются гармоническими.
В экосистеме возникает то, что называется биоритмом.
В заключении приведем график, построенный на основании отчетов по отстрелу зайцев и рысей в Канаде в течении столетия. Видно, что основные выводы, полученные в упрощенной схеме «хищник-жертва», справедливы и для этой реальной ситуации.

Дата добавления: 2018-04-05; просмотров: 620; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!