КОНЕЧНЫЙ МОЗГ. БОРОЗДЫ, ДОЛИ, ИЗВИЛИНЫ
ЛОКАЛИЗАЦИЯ ЦЕНТРОВ В КОРЕ ПОЛУШАРИЙ БОЛЬШОГО МОЗГА.
Конечный мозг развивается из передней части переднего мозгового пузыря и его полостью являются боковые желудочки. На сагитальном разрезе различают следующие поверхности полушарий: верхнелатеральную, нижнюю, и медиальную. Постоянные борозды (латеральная, центральная и теменно-затылочная) разделяют каждое полушарие на доли: лобную, теменную, затылочную и височную. В каждой доле имются борозды, извилины и дольки.На препаратах мозга на целом и сагиттальном разрезах определяется топография ядер следующих анализаторов (Рис. 8, 9): 1. Ядро двигательного анализатора, расположенное в передней центральной извилине и околоцентральной дольке. В двигательной зоне предцентральной извилины тело человека спроецировано вниз головой. При этом, правое полушарие получает сигналы от двигательного аппарата левой половины тела, а левое от правой. 2. Ядро двигательного анализатора артикуляции речи - в заднем отделе нижней лобной извилины, слева у правшей, справа у левшей.3. Ядро двигательного анализатора письменной речи в заднем отделе средней лобной извилины. При поражении этого анализатора наступает потеря способности писать. 4. Ядро кожного анализатора (болевой, температурной, тактильной чувствительности) - в постцентральной извилине и верхней теменной дольке. Тело человека спроецировано вниз головой. В связи с перекрестом чувствительных проводников в продолговатом и спинном мозге каждая постцентральная извилина получает сигналы с противоположной стороны тела. 5. Ядро зрительного анализатора письменной речи - в нижней теменной дольке (угловая извилина). При нарушении этого центра теряется способность читать (алексия). 6. Ядро слухового анализатора - на внутренней поверхности верхней височной извилины (извилина Гешля). При повреждении с обеих сторон теряется высший (корковый) анализ и синтез звуковых сигналов (корковая глухота). 7. Ядро слухового анализатора устной речи (центр Вернике) в задней части верхней височной извилины. С помощью этого анализатора человек контролирует свою речь, слышит и понимает чужую. При его выпадении теряется способность воспринимать слова, несмотря на сохранение слуха (словесная глухота). 8. Ядро обонятельного анализатора - в крючке парагиппокамповой извилины. При повреждении обонятельного анализатора больной не может различать запахи. 9. Ядро вкусового анализатора - в uncus gyri parahippocampalis. 10. Ядро зрительного анализатора - в затылочной доле по краям шторной борозды. При его одностороннем поражении, возникает половинная слепота (гемианопсия) на оба глаза.

Рис. 8.
1 - gyrus precentralis; 2 - gyrus frontalis inferior; 3 - gyrus frontalis medius; 4 - gyrus postcentralis; 5 - gyrus angularis; 6 - gyrus temporalis superior (извилина Гешля);7 - центр Вернике.

Рис. 9.
1 - gyrus precentralis; 2 - gyrus frontalis inferior; 3 - gyrus frontalis medius; 4 - gyrus postcentralis; 5 - gyrus angularis; 6 - gyrus temporalis superior (извилина Гешля);
7 – центр Вернике; 8,9 - uncus gyri parahippocampalis; 10 - sulcus calcarinus.
БАЗАЛЬНЫЕ ЯДРА ПОЛУШАРИЙ. БЕЛОЕ ВЕЩЕСТВО ПОЛУШАРИЙ. ВНУТРЕННЯЯ КАПСУЛА. БОКОВЫЕ ЖЕЛУДОЧКИ. ОБОНЯТЕЛЬНЫЙ МОЗГ.
На схеме горизонтального среза базальных ядер находятся хвостатое ядро, nucleus caudatus, чечевицеобразное ядро, nucleus lentiformis, оградку, claustrum и разделяющие их прослойки белого вещества: внутреннюю, наружную и внешнюю капсулы (рис. 10).
Особое внимание следует обратить на отделы (переднее бедро, crus anterior, колено, genus, и заднее бедро, crus posterior) внутренней капсулы и топографию проводящих путей:
1. В переднем отделе переднего бедра внутренней капсулы, ограниченном головкой хвостатого ядра и чечевицеобразным ядром, проходят волокна от переднего ядра зрительного бугра в кору лобной доли и обратные волокна из коры лобной доли к зрительному бугру, в заднем отделе - лобно-мостовой путь, tractus frontopontinus, идущий из коры лобной доли к собственным ядрам варолиевого моста своей стороны.
2. Через колено внутренней капсулы проходит корково-ядерный путь из коры нижней трети предцентральной извилины ко всем двигательным ядрам черепных нервов. 3. В передних двух третях заднего бедра внутренней капсулы проходит корково-спинномозговой путь, идущий из коры верхних двух третей предцентральной извилины к альфа-большим мотонейронам передних рогов серого вещества спинного мозга. Кзади от него проходит зрительнобугорно-корковый путь, tractus thalamocorticalis, от вентрального и латерального ядер зрительного бугра к коре постцентральный извилины и верхней теменной дольки. Далее через задний отдел заднего бедра внутренней капсулы последовательно проходят: затылочно-височный путь моста, tractus occipitotemporopontinus, и центральные слуховой и зрительный пути.
Хвостатое ядро и скорлупа чечевицеобразного ядра объединяются под названием полосатого тела, corpus striatum, являющегося высшим центром экстрапирамидной системы, которому подчинена паллидарная система, включающая бледный шар, globus pallidus, красное ядро, черное вещество и подталамическое (Люисово) тело. Кроме указанных ядер к экстрапирамидной системе относятся: нижняя олива, мозжечок, ретикулярная формация и зрительный бугор.
При изучении боковых желудочков необходимо обратить внимание на его отделы: передние, нижние, задние рога, центральный отдел и их стенки. Знать: полостью какой доли является тот или иной отдел и через что сообщается с полостью III желудочка. Необходимо иметь чёткое представление о местах образования цереброспинальной жидкости и путях её оттока.
Изучая обонятельный мозг, необходимо на препаратах рассмотреть его периферический (обонятельную долю) и центральные отделы. Периферический отдел представлен обонятельной луковицей, обонятельным трактом, обонятельным треугольником, передним продырявленным веществом и прозрачной перегородкой. К центральному отделу относят сводчатую извилину с крючком, uncus, в котором заложен корковый конец обонятельного анализатора; подмозолистую извилину; серое вещество, покрывающее верхнюю поверхность мозолистого тела и образующее две продольные полоски, striae longitudinalis, объединяющиеся позади утолщения мозолистого тела в так называемую серую повязку, fasciola cinerea, переходящую в зубчатую извилину, gyrus dentatus и, наконец, извилину морского коня (аммонов рог), hippocampus.
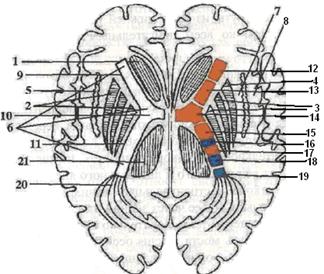
Рис.10.
1- nucleus caudatus; 2- nucleus lentiformis; 3- globus pallidus; 4- putamen; 5- claustrum; 6- capsula interna, 7- capsula externa; 8- capsula extrerna; 9- crus anterior capsula mterna; 10- genus capsula interna; 11- crus posterior capsula interna; 12- tractus frontothalamicus; 13- tractus frontopontinus; 14- tractus corticonuclearis; 15- tractus corticospinalis; 16- tractus thalamocorticalis; 17- tractus occipitotemporopontinus; 18- центральный слуховой путь; 19- центральный зрительный путь; 20- radiatio optica.
ДВИГАТЕЛЬНЫЙ АНАЛИЗАТОР
Двигательный анализатор осуществляет анализ и синтез раздражений, поступающих из двигательного аппарата, обеспечивает статику и динамику растущего организма ребенка, подвижность и трудовую деятельность человека.
Афферентный отдел его состоит из 3-х нейронов (рис. 11). Тела первых нейронов, псевдоуниполярных клеток, заложены в спинномозговых узлах и узлах V, IX и Х пар черепных нервов.
Дендриты первых нейронов, заложенных в спинномозговых узлах, направляются в составе периферических нервов к костям, суставам и мышцам, заканчиваясь в них рецепторами (проприорецепторами), воспринимающими раздражения, возникающие в связи с изменениями напряжения или растяжения мускулатуры, натяжения или расслабления сухожилий, суставных сумок и т.д. (мышечно-суставное чувство).
Воспринятое раздражение по дендритам передается телам клеток, аксоны которых в составе заднего корешка вступают в спинной мозг и, не прерываясь в сером веществе, поднимаются в заднем канатике. При этом аксоны первых нейронов, воспринимающих раздражение от мышц и суставов нижних конечностей и нижних отделов туловища, образуют нежные пучки, fasciculi gracilis, формирующиеся на протяжении 19 нижних сегментов спинного мозга.
Аксоны клеток, воспринимающих раздражения из мышц и суставов верхних конечностей, верхних отделов туловища и частично шеи, образуют расположенные латерально от предыдущих клиновидные пучки, fasciculi cuneati, формирующиеся на протяжении 12 верхних сегментов спинного мозга. Таким образом, ниже 4 грудного сегмента в заднем канатике проходит только пучок Голля, а выше - оба пучка (Голля и Бурдаха). Из спинного мозга эти пучки поднимаются в продолговатый мозг и заканчиваются в своих ядрах, nuclei fasciculi gracilis et cuneati. Клетки ядер нежного (Голля) и клиновидного (Бурдаха) пучков являются вторыми нейронами. Аксоны их идут двумя путями: меньшая часть, под названием наружных дугообразных волокон, следует через нижние ножки мозжечка к ядру шатра, nucleus fastigii, а большая часть их дугообразно огибает серое вещество, окружающее центральный канал (внутренние дугообразные волокна), переходит между оливами на противоположную сторону и поднимается по бокам от срединной линии кзади от пирамид, получая название медиальной петли, lemniscus medialis. При этом волокна ядер нежного пучка занимают более дорзальное положение. В мосту волокна медиальной петли проходят на границе основания и покрышки, пересекая поперечно идущие волокна трапециевидного тела. В верхнем отделе моста к ним присоединяется спиноталамический путь и весь комплекс волокон поднимается в покрышку ножек мозга, проходя кнаружи и дорзальнее красного ядра. На пути волокна медиальной петли отдают коллатерали к ядрам ретикулярной формации. Заканчивается медиальная петля в вентральном и латеральном ядрах зрительного бугра. Клетки вентрального и латерального ядер зрительного бугра являются третьими нейронами. Аксоны их, составляя таламокортикальный тракт, следуют через заднюю треть заднего бедра внутренней капсулы, позади пирамидного пути, в кору верхних двух третей передней центральной извилины, где заложена ядерная часть коркового конца двигательного анализатора. Эта часть афферентного отдела двигательного анализатора проводит проприоцептивную чувствительность от мышц, сухожилий, костей и суставов конечностей, туловища и частично шеи.



| Nucl.gracilis
(II нейрон)
| |


| Gangl.spinale Th5-Co1
(I нейрон)
| |

| Gangl.spinale
C1-Th4
(I нейрон)
| |

| Nucl. cuneatus
(II нейрон)
| |
| Nucl.tr.solitarii (II нейрон)
| |



| Gangl.rigeminale
(I нейрон)
| |




| Nucl.tr.mesencephalici n.trigemini (II нейрон)
| |




Рис. 11. Пути сомато-сенсороного анализатора
От мышц головы и мышц шеи висцерального происхождения проприоцептивная чувствительность проводится по черепным нервам. Первые нейроны этой части пути двигательного анализатора заложены в узлах V, IX и Х пар черепных нервов. Аксоны их следуют в составе нервов к мезенцефалическому ядру тройничного нерва, nucleus tractus mesencephali nervi trigemeni, и ядру одиночного пучка, nucleus tractus solitarii.
В этих ядрах заложены тела вторых нейронов. Аксоны вторых нейронов переходят на противоположную сторону и присоединяются к медиальной петле, достигая медиального и заднего отделов вентрального ядра зрительного бугра. Клетки этого ядра являются третьими нейронами, аксоны которых следуют в составе таламокортикального тракта в кору нижней трети передней центральной извилины. Таким образом, медиальная петля есть комплекс аксонов клеток ядер нежного и клиновидного пучков, собственного ядра заднего рога, желатинозной субстанции, а также чувствительных ядер тройничного, языкоглоточного и блуждающего нервов. Получив свое начало в продолговатом мозге, она заканчивается в зрительном бугре, являясь проводником общей чувствительности. Необходимо отметить, что часть аксонов первых нейронов (клеток узлов V, IX и Х пар черепных нервов) без перерыва в ядрах стволовой части мозга достигают коры передней центральной извилины. Таким образом, первые нейроны пути двигательного анализатора заложены в спинномозговых узлах и узлах V IX и Х пар черепных нервов, вторые нейроны — в ядрах нежного и клиновидного пучков, а также в чувствительных ядрах тройничного, языкоглоточного и блуждающего нервов, а третьи нейроны — в вентральном и латеральном ядрах зрительного бугра.
Поражение афферентной части двигательного анализатора ведет к выпадению мышечно-суставной (проприоцептивной) чувствительности, что выражается в нарушении представления о положении частей тела в пространстве, ощущения позы и производимых движений, нарушается координация движений (сенсорная атаксия). Больной с закрытыми глазами не может определить положение конечности или её частей, направления и объёма движений в том или ином суставе, движения теряют свою плавность, становятся неправильными, неточными.
Переработанная и проверенная в результате аналитико-синтетической деятельности корковых нейронов информация передается гигантским пирамидным клеткам Беца и пирамидным клеткам V и VI слоев коры, которые дают начало исполнительной системе, обеспечивающей движение импульсов из коры к мотонейронам спинного мозга и двигательным ядрам черепных нервов для осуществления рабочих, исполнительных актов, т. е. ответной реакции на воспринятые раздражения. Эта исполнительная система представлена пирамидным и экстрапирамидными путями или системами.
Пирамидная система осуществляет произвольные, высокодифференцированные движения, посылая из коры полушарий большого мозга через двигательные ядра черепных нервов и α-мотонейроны передних рогов спинного мозга импульсы к мышцам. Она имеет двух- или трехнейронный тип строения и представлена центральными (корковыми), промежуточными (возбуждающими или тормозными) и периферическими альфа-мотонейронами (Рис.12)..
Центральными (первыми) нейронами являются гигантские пирамидные клетки Беца (140 мкм) и большие пирамидные клетки пятого слоя коры передней центральной извилины, gyrus precentralis, лобной доли и околоцентральной дольки, lobulus paracentralis, а также большие и средние пирамидные клетки лобной, теменной, затылочной и височной долей.
Аксоны центральных нейронов, расположенных в пятом слое коры передней центральной извилины и околоцентральной дольки, идут более компактно, нежели аксоны клеток коры других долей. В связи с этим в пирамидной системе выделяют концентрированно расположенные и рассеянные волокна.
Концентрированно расположенные волокна осуществляют целенаправленную ответную реакцию на раздражения, полученные по афферентной части двигательного анализатора, а рассеянные волокна, по-видимому, обеспечивают ответную целенаправленную реакцию на раздражения, полученные по афферентным путям других анализаторов.
Возникнув в различных отделах коры, все эти волокна образуют лучистый венец и, постепенно сближаясь, собираются в компактный пучок, проходящий через колено и передние две трети заднего бедра внутренней капсулы.
В зависимости от ядер, к которым они направляются, в пирамидной системе различают корково-ядерный и корково-спинномозговой пути (рис. 12).
Корково-ядерный путь, tractus corticonuclearis, связывает кору большого мозга со всеми двигательными ядрами черепных нервов. Первыми нейронами этого пути являются гигантские и большие пирамидные клетки пятого слоя коры нижней трети передней центральной извилины. Аксоны их, постепенно сближаясь, образуют компактный пучок, который проходит через колено внутренней капсулы в основание ножки мозга. Здесь он, располагаясь латеральнее от лобно-мостового пути, tractus frontopontinus, отдает часть волокон к ядрам глазодвигательного, nucleus nervi oculomotorii, и блокового, nucleus nervi trochlearii, нервов своей и противоположной стороны. Из основания ножки мозга этот путь спускается в основание моста, отдавая часть волокон к двигательным ядрам тройничного, nucleus motorius nervi trigemini, отводящего, nucleus nervi abducentis, и верхней части ядра лицевого, nucleus nervi facialis, нервов своей и противоположной стороны, к нижней части ядра лицевого нерва только противоположной стороны и коллатерали к собственным ядрам моста, nuclei proprii pontis. Оставшаяся сравнительно небольшая часть волокон спускается в продолговатый мозг и заканчивается в двигательных ядрах IX, Х и XI пар черепных нервов своей и противоположной стороны, в ядре XII пары противоположной стороны. Необходимо подчеркнуть, что не все аксоны первых нейронов вступают в контакт непосредственно с клетками двигательных ядер. Большая часть их образует синаптическую связь с промежуточными нейронами - клетками ретикулярной формации, а последние, в свою очередь, - с клетками двигательных ядер.
Клетки двигательных ядер черепных нервов являются вторыми или третьими нейронами корково-ядерного пути. Аксоны их направляются в составе соответствующих нервов к мышцам головы и частично шеи, а также к мышцам глотки и гортани. Так, в составе глазодвигательного нерва - к внутренней, верхней, нижней прямым и нижней косой мышцам глаза, а также мышце, поднимающей верхнее веко; в составе блокового нерва - к верхней косой, а отводящего - к латеральной прямой мышцам глаза. Аксоны клеток двигательного ядра тройничного нерва идут в составе его третьей ветви ко всем жевательным, челюстно-подъязычной мышце, к мышцам, напрягающим мягкое небо и барабанную перепонку, и переднему брюшку двубрюшной мышцы. В составе лицевого нерва волокна идут к мимическим, шилоподъязычной мышцам и подкожной мышце шеи, а также к мышце стремени и заднему брюшку двубрюшной мышцы. В составе остальных нервов: IX- к шилоглоточной мышце; Х - к мышцам глотки, мягкого неба, гортани и верхней части пищевода; XI- к трапециевидной и грудино-ключично-сосцевидной мышцам; XII- к мышцам языка и мышцам, лежащим ниже подъязычной кости.


| Nucl.n.glosso-pharyngeus
et vagus
| |






| Tr. corticospinalis anterior
| |







| Tr. Corticospinalis lateralis
| |

Рис. 12. Пирамидные пути.
В связи с тем, что большинство двигательных ядер черепных нервов (III, IV, V, VI, IX, X, XI пар и верхней части ядра лицевого нерва) получают волокна из коры обоих полушарий большого мозга, одностороннее поражение первых (центральных) нейронов корково-ядерного пути ведет лишь к ограничению произвольных движений, снижению силы мышц, иннервируемых этими нервами, т.е. неполной утрате движений, называемой парезом. Поражение же первых нейронов, аксоны которых следуют к ядру подъязычного и нижней части ядра лицевого нервов из коры только противоположного полушария, ведет к возникновению центрального (спастического) паралича мышц языка и мимических мышц нижнего отдела лица на стороне, противоположной очагу поражения.
Центральный паралич характеризуется резким повышением тонуса мышц при пассивном движении (мышечная гипертония) и сухожильных рефлексов (гиперрефлексия), появлением сопутствующих движений (синкинезия) и патологических рефлексов. Развитие этих симптомов объясняется тем, что при поражении центральных (корковых) нейронов, оказывающих в обычных условиях тормозящее влияние на периферические нейроны, сохраняется целостность рефлекторных дуг, по которым периферические нейроны (клетки двигательных ядер) непрерывно получают раздражения с периферии и, в свою очередь, посылают импульсы к мышцам, повышая их тонус (мышцы напряжены, плотны на ощупь), усиливая сухожильные рефлексы, вызывая появление патологических рефлексов.
Поражение периферических нейронов корково-ядерного пути ведет к возникновению периферического, вялого, атрофического паралича мышц на своей стороне, характеризующегося, вследствие перерыва рефлекторной дуги, отсутствием рефлексов и тонуса мышц с последующей их дегенеративной атрофией.
Корково-спинномозговой путь, tractus corticospinalis, связывает кору больших полушарий с двигательными ядрами передних рогов спинного мозга. Первыми нейронами его являются гигантские клетки и большие пирамидные клетки коры верхних двух третей передней центральной извилины и околоцентральной дольки. Аксоны их следуют в составе лучистого венца и в области внутренней капсулы образуют компактный пучок, занимающий передние две трети ее заднего бедра. Из внутренней капсулы волокна корково-спинномозгового пути спускаются в основание ножек мозга, располагаясь латеральнее от корково-ядерного пути. Здесь весь пирамидный путь занимает центральный отдел основания ножки мозга, составляя 3/5 её. Далее этот путь спускается в основание моста, где поперечными волокнами, возникающими из его собственных ядер и идущими в мозжечок, разбивается на изолированные пучки, хорошо видимые на поперечных срезах моста. Здесь от пирамидных волокон отходят коллатерали к собственным ядрам моста, аксоны клеток которых переходят на противоположную сторону и в составе средних ножек мозжечка следуют к коре полушарий мозжечка .
В продолговатом мозге волокна корково-спинномозгового пути вновь собираются в компактные пучки, расположенные по бокам передней срединной щели - пирамиды. От них отделяется небольшой пучок волокон, идущих через нижние ножки в кору полушарий мозжечка (рис. 13).
Эти волокна вместе с коллатералями, отходящими от пирамидного пути к собственным ядрам моста, обеспечивают мозжечковую поправку к основному импульсу, идущему по пирамидному пути, заключающуюся в преодолении инерции движения.
На границе со спинным мозгом большая часть (80%) волокон пирамид переходит на противоположную сторону и спускается в боковом канатике спинного мозга под названием бокового корково-спинномозгового пути, tractus corticospinalis lateralis, посегментно заканчиваясь в двигательных ядрах передних рогов той же стороны.



Рис. 13. 1 - Пирамидный путь. 2 - Пучок волокон к мозжечку.
Меньшая часть (20%) волокон, под именем переднего корково-спинномозгового пути, tractus corticospinalis anterior, спускается в переднем канатике своей стороны до нижних грудных сегментов, посегментно заканчиваясь в двигательных ядрах передних рогов своей и противоположной стороны. Здесь волокна обоих путей вступают в контакт (синапс) с α-мотонейронами непосредственно, или через посредство промежуточных нейронов (интернейронов). Таким образом, α-мотонейроны являются вторыми или третьими нейронами корково-спинномозгового пути. Аксоны их выходят из спинного мозга через передние корешки и в составе ветвей спинномозговых нервов достигают мышц.
Одностороннее поражение центральных (первых) нейронов корково-спинномозгового пути ведёт к возникновению центрального, спастического паралича мышц конечностей противоположной стороны, при сохранении или ослаблении функции мышц шеи, туловища и промежности. Последнее объясняется тем, что эти мышцы получают двухстороннюю корковую иннервацию. Поражение пирамидного пути в спинном мозге выше шейного утолщения вызывает паралич мышц верхней и нижней конечностей одноименной стороны, а ниже — мышц нижней конечности той же стороны.
Поражение периферических двигательных нейронов (клеток передних рогов и их аксонов) ведет к возникновению периферического, вялого или атрофического паралича мышц той же стороны, характеризующегося утратой рефлексов, потерей тонуса мышц с последующей их дегенеративной атрофией.
ЭКТРАПИРАМИДНАЯ СИСТЕМА
Как корковые концы анализаторов не могут осуществлять точный анализ и синтез без одновременного влияния ретикулярной формации, так и пирамидный путь не может осуществлять точных и тонких движений без одновременного влияния экстрапирамидной системы.
Пирамидный путь обеспечивает произвольное сокращение той или иной мышцы, но даже самый примитивный двигательный акт требует согласованного участия многих мышц, последовательности их сокращений, а также различной силы и длительности сокращений. Это осуществляется экстрапирамидной системой.
У высших животных и человека экстрапирамидная система несет миостатическую функцию, создавая предуготованность мышц для выполнения волевых движений, осуществляемых пирамидным путем. Миостатическая функция делает двигательный акт плавным, гибким, точно соразмеренным во времени и в пространстве, обеспечивая оптимальную позу тела и наиболее выгодное положение отдельных мышечных групп для выполнения данного движения. Она обеспечивает реализацию заученных, стереотипных автоматизированных движений и регулирует тонус мышц. О роли экстрапирамидной системы в осуществлении двигательного акта может дать некоторое представление образное сравнение конечного двигательного пути с клавиатурой рояля, а мышц -со струнами, звучание которых зависит от сигнала, поступающего от клавиши. Стриопаллидарная система - это руки пианиста, нажимающие на клавиши двигательного пути с определенной последовательностью, силой и длительностью, создавая законченную мелодию стройного двигательного акта.
К экстрапирамидной системе относятся: стриопаллидарная система, нижняя олива, мозжечок, ретикулярная формация и зрительный бугор.
Стриопаллидарная система по морфологическим особенностям и функциональному значению разделяется на полосатое тело, corpus striatum, и палидум. Полосатое тело включает в себя хвостатое ядро, nucleus caudatus, и скорлупу, putamen, чечевичного ядра, являясь высшим центром экстрапирамидной системы. Паллидум, или паллидарная система, состоит из бледного шара, globus pallidus, красного ядра, nucleus ruber, черной субстанции, substantia nigra, и подталамического (люисова) тела, corpus subthalamicus. Высшей чувствительной инстанцией экстрапирамидной системы является зрительный бугор, thalamus.
Полосатое тело имеет двусторонние связи с корой лобной доли и зрительным бугром (рис. 14). Получив афферентные импульсы от двигательного аппарата через зрительный бугор, оно посылает сигналы в кору лобной доли, которая по корково-стриарным волокнам оказывает корригирующее влияние на его деятельность. В свою очередь, полосатое тело через подчиненные ему центры (бледный шар, красное ядро, подталамическое тело, черное вещество, ретикулярная формация и нижняя олива) оказывает влияние на альфа-малые мотонейроны передних рогов спинного мозга и клетки двигательных ядер черепных нервов, автоматически регулируя работу мышц и перераспределяя их тонус при движениях. Эта функция полосатого тела осуществляется по сложной системе нисходящих путей (рис. 14).
Далее волокна от бледного шара направляются к красному ядру, черному веществу, подталамическому телу, ядрам гипоталамуса, зрительному бугру и ядрам сетчатой формации. Из этих центров ведущее место занимает красное ядро, являющееся основным координационным центром экстрапирамидной системы. Получив импульсы от бледного шара, оно посылает их альфа-малым мотонейроиам переднего рога спинного мозга по красноядерно-спинальному пути, tractus rubrospinalis. Последний начинается от задней крупноклеточной части ядра и состоит из двух нейронов. Аксоны первых нейронов переходят на противоположную сторону, образуя вентральный перекрест покрышки ножек мозга (перекрест Фореля), и спускаются через ретикулярную формацию моста и продолговатого мозга в спинной мозг. Здесь они проходят в боковом канатике впереди от пирамидного пути и посегментно заканчиваются у альфа-малых мотонейронов переднего рога.
| Рис. 14. Экстрапирамидные пути
| |





















| Tr.thalamo-
rubroolivaris
| |

Последние являются вторыми нейронами. Аксоны их покидают мозг через передние корешки и в составе периферических нервов подходят к мышцам. Красное ядро оказывает влияние на альфа-малые мотонейроны и мышцы также через ретикулярную формацию и ядро нижней оливы. В первом случае аксоны клеток красного ядра в составе руброретикулярного пути переходят на противоположную сторону и заканчиваются в ядрах ретикулярной формации. Отсюда начинается ретикулоспинальный путь, спускающийся в боковом и переднем канатиках спинного мозга к альфа-малым мотонейронам переднего рога, аксоны которых следуют к мышцам. В другом случае аксоны клеток красного ядра и зрительного бугра в составе центрального пути покрышки (thalamorubroolivaris) следуют к ядру нижней оливы. От ядра нижней оливы часть волокон направляется через нижние ножки в кору полушарий мозжечка, обеспечивая связь полосатого тела с мозжечком. Большая же часть волокон в составе оливоспинального пути, tractus olivospinalis, спускается на границе переднего и бокового канатиков спинного мозга к альфа-малым мотонейронам переднего рога, от которых импульсы передаются к мышцам. Красное ядро, кроме того, имеет связи со зрительным бугром, а через него - с полосатым телом и корой полушарий большого мозга, а также прямые связи с корой. Связи красного ядра с мозжечком будут рассмотрены при описании последнего.
Черное вещество осуществляет регуляцию потока к мышцам тонических сигналов, посылаемых ретикулярной формацией. Получив импульсы от бледного шара и коры центральных и лобных извилин, оно передает их ядрам ретикулярной формации, красному ядру, верхним бугоркам четверохолмия, а через них - периферическим нейронам и мышцам, Кроме того, клетки черного вещества посылают волокна к зрительному бугру и ядрам глазных мышц.
Поражение черной субстанции вызывает нарушение пластического тонуса мышц в результате бесконтрольного прохождения к ним избыточных тонических сигналов, приводящих к резкому повышению их тонуса (ригидности),
Поражение полосатого тела, оказывающего тормозящее влияние на паллидарную систему, приводит к появлению избыточных непроизвольных движений (гиперкинез) и понижению мышечного тонуса (гипотония).
Поражение бледного шара проявляется бедностью и маловыразительностью движений (олигокинезия), их замедленностью, тонус мышц повышен. Больные малоподвижны, инертны, скованны, движения в суставах становятся толчкообразными (симптом зубчатого колеса) и др.
Поражение подталамического (Люисова) тела ведет к появлению размашистых движений в суставах конечностей. Важнейшим центром экстрапирамидной системы, осуществляющим регуляцию тонуса мышц и координацию движений, является мозжечок. Координация движений требует постоянной всесторонней информации от двигательного и вестибулярного аппаратов о положении тела в пространстве. Эта информация поступает в мозжечок по трем парам ножек.
Основным каналом поступления в мозжечок афферентных сигналов являются его нижние ножки. В составе их проходят волокна к ядру шатра, nucleus fastigii, и в кору мозжечка. К ядру шатра идут наружные дугообразные волокна от ядер нежного (Голля) и клиновидного (Бурдаха) пучков, волокна от ядер вестибулярного нерва (главным образом от ядер Бехтерева) и от ядер ретикулярной формации. Получив разрозненную информацию из различных источников, ядро шатра посылает её клеткам Пуркинье коры мозжечка, распределяя соответственно соматотопической проекции тела: в передние отделы коры червя - от головы и шеи, а в задние — от туловища; в передние отделы полушарий - от рук, а в задние - от ног, причем, проксимальные отделы конечностей проецируются медиальнее, чем дистальные (рис. 15).
Таким образом, полушария ответственны за координацию движений конечностей, а червь — головы, шеи и туловища. Ядро шатра, получив и переработав информацию, посылает её не только клеткам Пуркинье коры мозжечка. Часть волокон идёт в обратном направлении по проходящим через нижние ножки мозжечково-ретикулярному и мозжечково-вестибулярному путям к ядрам ретикулярной формации и вестибулярному ядру Дейтерса.
От ядер ретикулярной формации импульсы передаются по ретикулоспинальному пути, tractus reticulospinalis, альфа-малым мотонейронам спинного мозга и мышцам. Ядро Дейтерса, получив сигналы от ядра шатра, передает их альфа-малым мотонейронам спинного мозга по вестибулоспинальному пути, tractus vestibulospinalis. Волокна последнего, начавшись от ядра Дейтерса, спускаются через продолговатый мозг в передний канатик спинного мозга и посегментно заканчиваются синапсами с альфа-малыми мотонейронами переднего рога. Альфа-малые мотонейроны являются вторыми нейронами этого пути. Аксоны их в составе передних корешков и ветвей спинномозговых нервов следуют к мышцам.
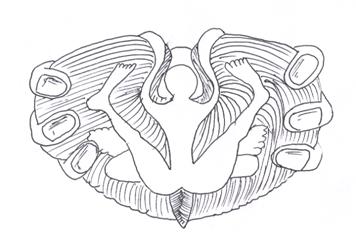
Рис. 15. Соматотопическая проекция тела на кору мозжечка
В кору мозжечка через нижнюю ножку идут отмеченные выше волокна от пирамидного пути и ядра нижней оливы, а также задний спинно-мозжечковый путь Флексига, tractus spinocerebellaris posterior, (рис. 16). Первыми нейронами последнего являются псевдоуниполярные клетки межпозвоночных узлов. Периферические отростки (дендриты) в составе нервов следуют к мышцам, заканчиваясь в них мышечными веретенами (проприорецепторами). Центральные отростки (аксоны) в составе задних корешков идут к грудному ядру основания заднего рога, nucleus thoracicus. Здесь лежат тела вторых нейронов, аксоны которых направляются в боковой канатик своей стороны и образуют пучок, занимающий его заднюю периферию. Волокна этого пути поднимаются в продолговатый мозг и по нижней ножке мозжечка следуют к коре верхнего червя. Таким образом, через нижние ножки мозжечка проходят как афферентные (волокна от пирамидного пути, ядер нежного и клиновидного пучков, задний спинно-мозжечковый, оливо-мозжечковый, вестибуло-мозжечковый, ретикуло-мозжечковый), так и эфферентные (мозжечково-вестибулярный, мозжечково-ретикулярный) пути.
| Cпинномозговой узел
( I нейрон)
| |


| Tr. spinocerebellaris post.
| |
| Tr.spinocerebellaris ant.
| |







| Tr.spinocerebellaris ant.
| |
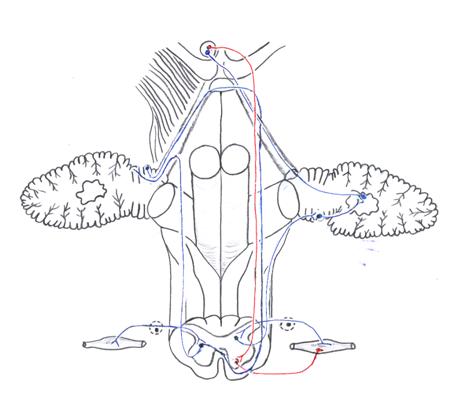
Рис. 16. Спинномозжечковые пути
Проприоцептивная чувствительность проводится в мозжечок и по переднему спинно-мозжечковому пути Говерса, tractus spinocerebellaris anterior(рис. 16). Тела первых нейронов его лежат в межпозвоночных узлах. Периферические отростки (дендриты) их заканчиваются рецепторами (проприорецепторами) в мышцах, сухожилиях, надкостнице, связках и суставах. Центральные отростки (аксоны) в составе задних корешков вступают в спинной мозг и заканчиваются в промежуточно-медиальном ядре боковой промежуточной зоны. Здесь лежат тела вторых нейронов. Аксоны их идут двумя путями: часть следует в боковой канатик своей, а часть — переходит через переднюю белую спайку в боковой канатик другой стороны и образует компактный пучок, занимающий переднюю периферию канатика. Из спинного мозга все эти волокна поднимаются через продолговатый мозг, дорзальную часть моста (кнаружи от медиальной петля) в верхний мозговой парус, где волокна, перешедшие в спинном мозге на противоположную сторону, образуют второй перекрест и возвращаются на свою сторону. Далее они следуют в составе верхней ножки мозжечка к коре нижнего червя.
Особым афферентным путем мозжечка, корригирующим его деятельность, является двухнейронный корково-мосто-мозжечковый путь (рис. 17). Тела первых нейронов его заложены в коре лобной, теменной, затылочной и височной долей полушарий большого мозга. В зависимости от локализации тел первых нейронов и особенностей хода их аксонов до собственных ядер моста, в нем различают лобно-мостовой, tractus frontopontinus, и теменно-затылочно-височно-мостовой, tractus occipitotemporopontinus, пути.
Лобно-мостовой путь образуется аксонами клеток коры передних отделов верхней и средней лобных извилин, спускается через задний отдел переднего бедра внутренней капсулы, медиальный отдел основания ножки мозга, занимая её 1/5 часть, и заканчивается в собственных ядрах моста своей стороны.
Теменно-затылочно-височно-мостовой путь образуется аксонами клеток коры теменной, затылочной и височной долей. Он проходит через задний отдел заднего бедра внутренней капсулы позади таламо- кортикального тракта, в наружном отделе основания ножки мозга, занимая 1/5 часть, и заканчивается в собственных ядрах моста своей стороны.
В собственных ядрах моста заложены тела вторых нейронов корково-мосто-мозжечкового пути. Их аксоны переходят на противоположную сторону и в составе средних ножек мозжечка, под названием мосто-мозжечкового пути, следуют к коре полушарий мозжечка. Таким образом, полушария большого мозга связаны с противоположными полушариями мозжечка. Поэтому при поражении коры больших полушарий расстройства координации возникают на противоположной очагу стороне.
Полученная по всем афферентным путям и переработанная клетками Пуркинье информация передается зубчатому ядру. Аксоны клеток этого ядра в составе верхних ножек мозжечка идут к красному ядру и зрительному бугру противоположной стороны. Перекрест волокон происходит на уровне нижнего двухолмия. Аксоны клеток красного ядра дают начало руброспинальному пути, который образует вентральный перекрест покрышки ножек мозга и следует к альфа-малым мотонейронам переднего рога спинного мозга. От последних импульсы передаются мышцам. По волокнам же, следующим к зрительному бугру, передается информация его клеткам, а от них - коре лобной доли и стриопаллидуму. Таким путем мозжечок связан с корой и стриопаллидарной системой.
Функция мозжечка заключается в рефлекторном поддержании мышечного тонуса, равновесия, координации и синергии движений, а также трофики тканей. Поэтому поражение мозжечка, а также афферентных и эфферентных его путей вызывает нарушение равновесия тела при стоянии (статическая атаксия) и ходьбе (динамическая атаксия), связанное с нарушением согласованности мышц-антагонистов (асинергия). Эти расстройства сопровождаются резким снижением мышечного тонуса (гипотония). Мышцы становятся дряблыми, вялыми, увеличивается объем движений в суставах. Согласно соматотопической проекции в коре мозжечка поражение червя ведет к грубым нарушениям статики вследствие утраты синергии мышц, стабилизирующих центр тяжести туловища, а поражение полушарий мозжечка – к возникновению атаксии конечностей на стороне поражения. Последнее объясняется тем, что волокна от зубчатого ядра мозжечка к красному (дентато-рубральный тракт) и волокна руброспинального пути совершают перекресты.
КОЖНЫЙ АНАЛИЗАТОР
Кожный покров тела (кожа), достигающий у взрослых площади около 1,6 квадратных метра, находится под постоянным воздействием окружающей среды, непрерывно получая из нее огромное количество раздражений (температурные, тактильные, болевые и др.). Анализ и синтез этих раздражений осуществляется кожным анализатором. Его афферентная часть представлена проводящими путями болевой, температурной, тактильной, а также пространственной кожной чувствительности. Болевая и температурная чувствительности от кожи шеи, туловища и конечностей проводятся спинномозговыми нервами, а от кожи головы – тройничным, блуждающим и верхними шейными спинномозговыми нервами (рис. 18).
Первыми нейронами, воспринимающими болевую и температурную чувствительность от кожи шеи, туловища и конечностей, являются ложно-униполярные клетки спинномозговых узлов. Их периферические отростки (дендриты) идут в составе спинномозговых нервов к коже, заканчиваясь рецепторами, а центральные отростки (аксоны) следуют в составе заднего корешка спинномозгового нерва к телам вторых нейронов, которые располагаются в собственном ядре заднего рога.





| Кора затылочной и височной долей
| |
| Tr.occipito-
temporo-
pontinus
| |



| Crus posterior capsula inerna
| |

| Crus anterior capsula inerna
| |


Рис. 17. Корково-мосто-мозжечковые пути
Аксоны вторых нейронов идут через переднюю белую спайку на противоположную сторону и образуют пучок, поднимающийся в боковом канатике медиальнее от переднего спинно-мозжечкового пути под названием бокового спинно-таламического тракта. При этом в передней части его располагаются волокна, проводящие болевую чувствительность, а в задней - температурную. Из спинного мозга волокна этого пути следуют через продолговатый мозг в покрышку моста, где присоединяются к медиальной петле и в ее составе достигают вентрального и латерального ядер зрительного бугра. Здесь заложены тела третьих нейронов. Аксоны третьих нейронов проходят позади пирамидного пути через заднее бедро внутренней капсулы в составе таламокортикального пути к коре верхних двух третей задней центральной (постцентральной) извилины, в которой заложено ядро коркового конца кожного анализатора.
Первыми нейронами, воспринимающими болевую и температурную чувствительность от кожи головы, являются ложно-униполярные клетки полулунного узла тройничного нерва, верхнего узла блуждающего и узлов трех верхних шейных спинномозговых нервов.
Периферические отростки (дендриты) следуют в составе ветвей тройничного нерва к коже лица, ушной раковины и лба, в составе ушной ветви блуждающего и большого ушного нервов — к коже наружного слухового прохода и ушной раковины, а также в составе большого и малого затылочных нервов – к коже затылочной области.
Центральные отростки (аксоны) клеток узла тройничного нерва по его чувствительному корешку подходят к главному чувствительному ядру, nucleus sensorius principalis, и ядру спинального тракта; аксоны клеток верхнего узла блуждающего нерва – к ядру одиночного пучка, nucleus tractus solitarii, а аксоны клеток верхних шейных узлов – к собственному ядру заднего рога. В этих ядрах лежат тела вторых нейронов. Аксоны их, перейдя на противоположную сторону, присоединяются к медиальной петле и в её составе достигают латерального ядра зрительного бугра. Здесь заложены тела третьих нейронов, аксоны которых в составе таламокортикального тракта следуют к коре нижней трети задней центральной извилины.
В связи с тем, что аксоны вторых нейронов пути болевой и температурной чувствительности переходят на противоположную сторону, при поражении их слева выпадает чувствительность на правой стороне тела.
| Рис. 18. Пути болевой и температурной чувствительности
| |

| Nucl. proprius
cornu
posterior
(II нейрон)
| |
| Tr. spinothalamicus lateralis
| |


| Gangl.superior
(X пара) ( I нейрон)
| |

| Nucl. tr.solitarii
(II нейрон)
| |


| Nucl.sensorius principalis et nucl.tr.spinalis (V пара) (III нейрон)
| |






Дата добавления: 2018-02-28; просмотров: 515; | Поделиться с друзьями:
|

Мы поможем в написании ваших работ!