Лесной биотический насос и естественный отбор
Заселение суши жизнью в геологическом прошлом путем увлажнения почвы, на которой могут произрастать растения, является, как мы видели, сложной биологической проблемой. В русском языке слова суша и сушь однокоренные, что подчеркивает сложность поддержания запасов влаги на суше. Суша, лишенная жизни, представляла собой сухую пустыню, над которой не было испарения, и океанская влага не могла поступать на сушу даже в прибрежной полосе. Какая растительность была пионером заселения суши, мы не знаем. На этот вопрос могут дать ответ лишь кропотливые палеонтологические исследования. Вероятно, это были древние травы и кустарники, которые смогли организовать муссонный климат в прибрежной полосе.
С эволюционным открытием возможностей лесного покрова, первичные леса начали продвигаться все дальше вглубь континентов, образуя речные бассейны. Совершенствование биотического лесного насоса привело к эволюционному образованию современных деревьев во всех климатических зонах. Леса, образованные этими деревьями, поддерживали оптимальную влажность почвы на любом удалении от океана при любых уклонах суши, включая все горные области. Леса достигли наибольшей продуктивности, ограничиваемой только потоком солнечной энергии. Пионерная растительность, заселявшая сушу, была вытеснена лесами практически на всей территории континентов. Пустыни перестали существовать на суше.
|
|
|
Биотический насос влаги — это синхронное действие всех деревьев и обслуживающих деревья остальных организмов сообщества. Сложная физиология каждого вида живого организма возникла в результате эволюционного естественного отбора особей в многочисленных популяциях вида. Биотический насос влаги не менее сложное образование, чем физиология самых высокоорганизованных видов биосферы. Поэтому он мог возникнуть также только путем естественного отбора индивидуальных деревьев и других жестко связанных с деревьями видов сообщества. Биотический насос не может быть следствием примитивной физической самоорганизации, подобной, например, образованию циклонов и антициклонов в атмосфере.
Как же могла возникнуть скоррелированность действий всех деревьев в лесу, которая необходима для организации биотического насоса влаги, если деревья конкурентно взаимодействуют друг с другом? Естественный отбор направлен на закрепление преимуществ каждого отдельного дерева — сильные деревья закрепляются в популяции, а слабые вытесняются из нее. Хороший биотический насос дает преимущество всем деревьям в лесу. От плохого все одинаково страдают, и те, которые затрачивают свои жизненные силы на создание насоса, и те, которые ничего не делают для поддержания его работы. Эти последние могут использовать высвободившуюся энергию на вытеснение работающих на насос деревьев из популяции, разрушая действие всего лесного биотического насоса. Для многих биологов в мире это служит кажущимся логическим обоснованием невозможности существования биотического лесного насоса влаги (как и любого другого подобного биотического механизма регуляции окружающей среды). Он представляется противоречащим принципам естественного отбора.
|
|
|
Это противоречие снимается в общей концепции биотической регуляции окружающей среды (Горшков, 1995, www.biotic-regulation.pl.ru), учитывающей существование чувствительности или разрешающей способности e реакции организмов на любые изменения окружающей среды. Если относительные изменения параметров среды меньше e, организмы сообщества не реагируют на это изменение. Если изменение больше e, организмы начинают воздействовать на окружающую среду, стремясь вернуть ее в оптимальное состояние. Совершенно так же мы начинаем приводить свое жилище в порядок, когда накопившаяся грязь и мусор становятся для нас заметными.
|
|
|
В лесу деревья реагируют на локальную влажность почвы. Если влажность почвы вокруг дерева уменьшается или увеличивается на величину, большую e, относительно оптимального значения, дерево начинает увеличивать или уменьшать транспирацию листьев, изменять выбросы в атмосферу биологических ядер конденсации влаги и производить массу других действий, которые нам неизвестны благодаря их чрезвычайной сложности. Дерево, правильно воздействующее на окружающую среду, улучшит локальную влажность почвы, вернув ее к оптимальному состоянию. Деревья-мутанты с ухудшенной чувствительностью не будут производить воздействий на локальную окружающую среду. Это приведет к ухудшению среды вокруг них, потере ими конкурентоспособности и вытеснению их из популяции нормальных деревьев. В результате, в популяции сохраняются только нормальные деревья, которые все будут реагировать одинаково на изменение окружающей среды, что и создаст действие лесного биотического насоса. Таким образом, наблюдаемое существование лесного биотического насоса, описанное в предшествующих разделах, не противоречит естественному отбору и является одним из основных подтверждений существования общей биотической регуляции окружающей среды.
|
|
|
Заключение
Биотический насос атмосферной влаги работает в естественных ненарушенных лесах. Только естественные первичные леса, аборигенные для данной территории, способны обеспечить долгосрочное устойчивое функционирование биотического насоса влаги, так как именно их генетические свойства скоррелированы с геофизическими особенностями той местности, в которой они растут. Искусственные насаждения из экзотических деревьев с генетическими программами, не соответствующими данной местности, не способны неограниченно долго самоподдерживаться на чуждой им территории, не приводя к деградации условий окружающей среды. Вторичные леса, восстанавливающиеся после антропогенных нарушений или природных катастроф типа пожаров и ветровалов, находятся в процессе восстановления всех регулирующих окружающую среду функций, включая функцию биотического насоса, который поэтому в таких лесах сильно ослаблен.
Нормальное функционирование лесного биотического насоса речного бассейна возможно, если лесной покров непосредственно выходит на береговую границу с океаном или отстоит от океана на расстояние, много меньшее длины затухания геофизических потоков влаги с океана на сушу. Два крупнейших тропических речных бассейна Амазонки и Конго обладают лесным покровом дождевых лесов, непосредственно граничащих с береговой линией. Бассейны северных рек России, Канады и Аляски имеют лесной покров тайги, простирающийся до крайних северных пределов, где граничат с сильно заболоченными пространствами, выходящими на берег океана.
Уничтожение лесного покрова на береговой полосе шириной порядка длины затухания обрывает действие биотического насоса атмосферной влаги с океана. Лесной покров в глубине континента теряет способность компенсировать речной сток. Влага почвы стекает в океан, речной бассейн перестает существовать и леса засыхают. Весь запас влаги на суше стекает в океан за 4 года. Поэтому полное уничтожение лесного биотического насоса, т.е. сплошного покрова ненарушенного леса, переводит любой речной бассейн в пустыню за время порядка нескольких лет.
На Австралийском континенте покрытый лесом речной бассейн, совпадающий по площади с речным бассейном Амазонки, прекратил существование около 50-100 тысяч лет назад. Примерно в это же время Австралию начали осваивать люди. Существует множество косвенных доказательств того, что лесной покров Австралии был уничтожен аборигенами. Приведенное здесь рассмотрение объясняет механизм этого уничтожения.
После того, как появившиеся на материке люди в ходе хозяйственной деятельности или в результате пожаров уничтожили лесной покров по всему периметру Австралии на узкой береговой полосе порядка длины затухания, биотический насос, снабжавший водой внутренние части материка, оказался отрезанным от воды. В результате стока и испарения влаги почва высохла, и леса прекратили свое существование даже в отсутствие антропогенной активности и пожаров внутри континента. Кратковременность перехода в пустыню не оставила об этом процессе никаких следов в палеоданных. Практически все пустыни мира имеют непосредственный выход на берега океанов или внутренних морей. Поскольку береговая зона является предпочтительными при выборе людьми их поселений, появление всех протяженных пустынь на Земле является следствием деятельности человека.
Практика эксплуатации и вырубки лесов, ответственная за современную рекордную скорость прямого уничтожения лесов во всем мире, зародилась в Западной Европе, все области которой удалены от береговой границы океана и внутренних морей на расстояния, меньшие длины затухания геофизических потоков влаги с океана на сушу. Поэтому полное уничтожение естественных лесов в Западной Европе, заканчивающееся сейчас в Швеции и Финляндии, не привело к полному опустыниванию этих территорий, что до сих пор создает иллюзию возможности экспортировать практику уничтожения лесов в другие области планеты, для которых она является губительной. Отметим, что и в Западной Европе участились катастрофические наводнения, засухи и пожары, главным образом, за счет уничтожения оставшихся естественных лесов в горных районах. Эта практика уничтожения горных лесов, увеличивающих осадки в горах, привела также к резкому сокращению горных ледников, которое ошибочно приписывается предполагаемому глобальному потеплению, вызванному ростом атмосферного CO2.
Искусственные водохранилища, создаваемые в речных бассейнах с уничтожением естественных лесов на больших площадях, ликвидируют управляющее действие транспирации этих лесов, ослабляют действие биотического лесного насоса и, следовательно, уменьшают речной сток и осадки.
Изложенные результаты намечают возможную стратегию восстановления необходимого для жизни людей водного режима суши, который может привести к обводнению пустынь и засушливых территорий. Ликвидация лесного покрова в крупнейших речных бассейнах мира за времена порядка нескольких лет приведет к уменьшению речного стока на порядок величины, возникновению засух, наводнений и пожаров, частичному опустыниванию в прибрежной полосе и полному — внутри континентов. Экономические потери от этого на много порядков величины превысят экономические выгоды от вырубки лесов. Поэтому целесообразно срочно пересмотреть лесную политику во всем мире.
Прежде всего, необходимо немедленно прекратить освоение оставшихся ненарушенных лесов на всех территориях речных бассейнов, а не только на расстоянии нескольких километров вдоль крупных рек, и, в особенности, на территориях выхода лесов на берега океана и внутренних морей. Далее необходимо начать планомерное восстановление аборигенного лесного покрова на территориях, прилегающих к естественным лесам. Только на этих территориях восстановленные леса смогут устойчиво поддерживать водный режим и в дальнейшем самостоятельно расширять речной бассейн. Лесной покров может быть восстановлен на большей части не используемых в настоящее время засушливых территорий. Экологически грамотное человечество будущего навсегда получит в свое распоряжение неограниченные водные ресурсы восстановленных речных бассейнов.
Литература
Горшков В.Г., Макарьева А.М. (2006) Биотический насос атмосферной влаги, его связь с глобальной атмосферной циркуляцией и значение для круговорота воды на суше. Препринт № 2655, Петербургский институт ядерной физики, Гатчина, 49 с. www.biotic-regulation.pl.ru/offprint/vod_pr1.pdf
Горшков В.Г. (1995) Физические и биологические основы устойчивости жизни. ВИНИТИ, Москва, 470 с.
Данные, использованные в работе, относятся к Проекту Общественная Модель Климатической Системы (Community Climate System Model project), осуществляемому Директоратом Наук о Земле Национального Научного Фонда и Бюро Биологических и Экологических Исследований Департамента Энергии США. Данные распространяются Университетом Нью Гэмпшира (University of New Hampshire, EOS-WEBSTER Earth Science Information Partner (ESIP)) на сайте http://eos-webster.sr.unh.edu.

Рис. 1. Геофизические районы, где исследована зависимость среднегодовых осадков от расстояния до океана.

Рис. 2. Зависимость количества осадков P (мм/год) от расстояния x (км) от океана в обезлесенных территориях (полые символы) и территориях, покрытых лесом (черные символы). Нумерация районов как на рис. 1.
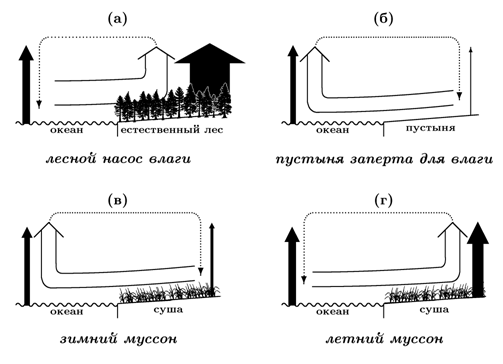
Рис. 3. Физический принцип распространения воздуха у земной поверхности из областей с меньшим испарением в области с большим испарением. Черные стрелки — потоки испарения, величина потока пропорциональна толщине стрелки. Полые стрелки — горизонтальные и восходящие потоки влажного воздуха. Пунктирные стрелки — горизонтальные и нисходящие потоки сухого воздуха.
Дата добавления: 2018-02-15; просмотров: 733; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!