Характеристика отрядов птиц: гагарообразные, поганкообразные, буревестникообразные.
Министерство сельского хозяйства Российской Федерации
Федеральное государственное бюджетное образовательное учреждение в ысшего образования
«Вятская государственная сельскохозяйственная академия»
БИОЛОГИЧЕСКИЙ ФАКУЛЬТЕТ
Кафедра Экологии и зоологии
КОНТРОЛЬНАЯ РАБОТА
ФИО Морозов А., форма обучения заочная
направление подготовки 06.03.01 Биология
направленность (профиль) программы Охотоведение
курс 2 , группа Бобз-112
Дисциплина: Зоология позвоночных
Тема: Вариант 20.
1. Миксины – паразиты рыб, особенности строения в связи с узкой специализацией.
2. Двоякодышащие. Особенности строения, биологии и распространения. Однолегочные, двулегочные двоякодышащие.
3. Происхождение и эволюция пресмыкающихся.
4. Характеристика отрядов птиц: гагарообразные, поганкообразные, буревестникообразные.
.
Руководитель: преподаватель Букина Л.А.
Регистрационный номер на кафедре
Дата регистрации «______»____________г.
Результат проверки _________________
Содержание
1. Миксины – паразиты рыб, особенности строения в связи с узкой специализацией.
2. Двоякодышащие. Особенности строения, биологии и распространения. Однолегочные, двулегочные двоякодышащие.
3. Происхождение и эволюция пресмыкающихся.
4. Характеристика отрядов птиц: гагарообразные, поганкообразные, буревестникообразные.
|
|
|
Микси́новые[1] (лат. Myxinidae) — семейство бесчелюстных, единственное в отряде миксинообразных[1] (Myxiniformes) и классе миксин[1] (Myxini). Относятся к самым примитивным современным черепным[2]. По состоянию на 2016 год известно 6 родов и 78 видов[1][1][1][2][3]
Подкласс Миксины (Myxini)

Миксины — паразиты, питающиеся как сонной, так и живой рыбой, в тело которой они глубоко внедряются. Таким образом, миксины являются хотя и временными, но настоящими эндопаразитами, представляя в этом отношении единственный пример среди позвоночных.
В связи с паразитическим образом жизни миксины во многих отношениях претерпели регрессивную эволюцию: глаза, спинной плавник и скелет жаберного аппарата у них атрофировались, а во внутреннем ухе сохранился лишь один полукружный канал. С другой стороны, они приобрели ряд приспособлений к паразитическому образу жизни: щупальца, расположенные по краям предротовой воронки, чрезвычайно мощный язык, вооруженный, как и у миног, роговыми зубцами и играющий роль сверлящего органа. У многих форм наружные жаберные отверстия не открываются на поверхности тела, а продолжаются в каналы, которые с каждой стороны впадают в общий парный канал, идущий под кожей и открывающийся наружу далеко сзади. Такое устройство дыхательных путей позволяет животному дышать, в то время как передняя часть его тела глубоко погружена в ткани жертвы. Половая железа молодых особей вырабатывает сперматозоиды, старых особей — яйца. Разновременное созревание половых продуктов исключает самооплодотворение.
|
|
|

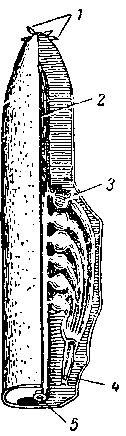
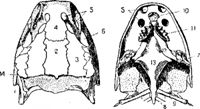
Миксина. I — цельное животное сбоку; II — головной конец снизу (по Гудричу):
1 — щупальцы, 2 — отверстия слизеотделительных полостей, 3 — левое и правое общие жаберные отверстия, 4 — ноздря, 5 — рот
С другой стороны, по строению кишечного тракта миксины отклонились от исходного типа слабее, чем миноги, так как обособленной дыхательной трубки у них нет, и глотка, прободенная жаберными отверстиями, непосредственно переходит в пищевод. В отличие от миног у миксин имеются большие слизеотделительные полости, которые расположены двумя продольными рядами по бокам тела и выделяют огромное количество слизи. Число жаберных мешков достигает пятнадцати, ноздря открывается на верхнем краю ротовой воронки, и гипофизарный вырост сообщается с глоткой. Последнее явление представляет, по-видимому, приспособление для дыхания: миксины часто зарываются в ил, выставив наружу лишь конец головы, и вода проходит через это отверстие в глотку и далее, в жаберные мешки.
|
|
|
Жаберный аппарат миксины (по Гудричу):
1 — щупальцы, 2 — глотка, 3 — жаберный мешок, 4 — общий жаберный канал, 5 — пищевод
Яйца миксин крупные, богатые желтком, развитие протекает без превращения.
Миксин насчитывают около 15 видов: все они чисто морские животные и в водах России не встречаются. Поедая рыбу, попавшуюся в сети, миксины местами приносят иногда значительный вред рыбоводству. Сами миксины промыслового значения не имеют.
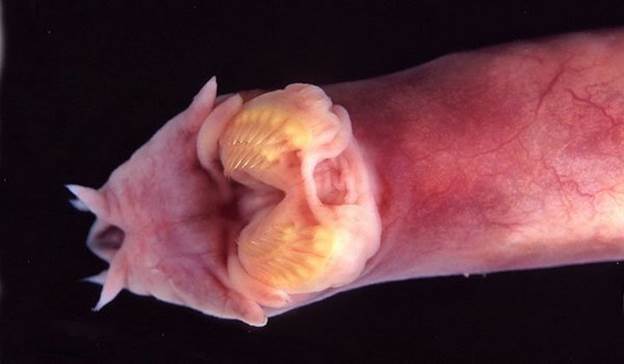
Миксина настолько скользкая, что человек не способен удержать её в руках, а рыбы-хищники не могут ее одолеть. Когда к миксине приближается враг, её поры начинают выделять слизь в огромном количестве, которой хищники могут подавиться. Когда опасность миновала, миксина сбрасывает обволакивающую её тело слизь и даже чихает, чтобы освободить от вязкого вещества свою единственную ноздрю. Эту ноздрю, а также рот миксины окружают мясистые усы. Круглый рот снабжен роговыми зубами: два ряда зубов расположены с каждой стороны языка, а один непарный зуб — на небе. У этой рыбы совсем не видно глаз — они затянуты кожей. Органами зрения служат светочувствительные клетки, расположенные на задней части червеобразного тела миксины.
|
|
|
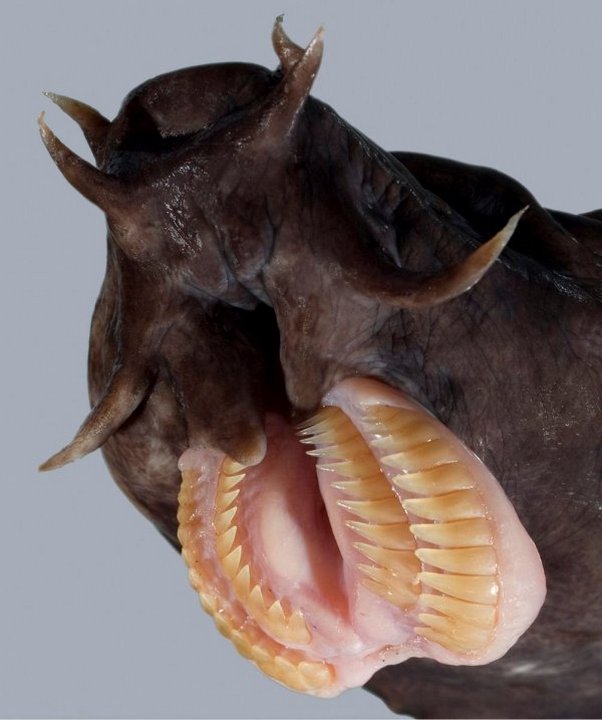
Слизь выполняет не только защитную функцию, но и помогает охотиться: миксины — хищники. Например, если жертвой миксины становится больная, но ещё способная сопротивляться рыба, то миксина выпускает под жаберную крышку своей добычи слизь, после чего жертва задыхается. Скользкое круглоротое животное охотится не только на пораженных болезнями и умирающих рыб, но и на вполне здоровых. Завладев рыбой, миксина прогрызает дыру в её теле и начинает поедать внутренности или высасывать мясо до костей.
Иногда миксины охотятся группами. Так, рыбаки Южной Норвегии, Англии и Западной Швеции часто обнаруживают у себя в сетях рыбу, которой лакомятся сразу несколько миксин одновременно. Чаще всего миксина предпочитает есть осетра, пикшу, треску, скумбрию и сельдь, но не откажется и от другой рыбы. Сама же миксина считается несъедобной.
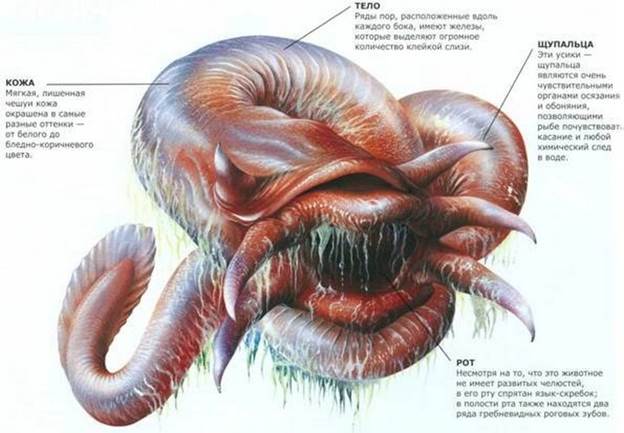
Миксина (лат. Myxini) обыкновенная обитает на глубинах 100-500 метров, преимущественный ареал обитания — около берегов Северной Америки, Европы, Исландии, Восточной Гренландии. Иногда можно встретить и в Адриатическом море. В зимнее время миксина иногда спускается и на большие глубины – вплоть до 1 км.
Размер этого животного невелик – 35-40 сантиметров, хотя иногда встречаются гигантские экземпляры – 79-80 сантиметров. Карл Линней, открывший это животное, первоначально даже включил его в класс червей из-за его специфичного внешнего вида. Хотя на самом деле миксины относятся к классу круглоротых, являющихся историческими предшественниками рыб. Окраска миксин может быть различной, однако, преимущественные цвета – розоватый и серо-красный.

Отличительной особенностью миксин является наличие ряда отверстий, выделяющих слизь, которые расположены вдоль нижнего края тела животного. Следует отметить, что слизь – это очень важный секрет миксин, который применяется животным для внедрения в полость рыбы, выбранной в качестве жертвы. Слизь имеет важную роль и в дыхании животного. Миксина – это настоящий завод по созданию слизи, в частности, если ее положить в ведро, полное воды, то уже через некоторое время вся вода преобразуется в слизь.
Плавники миксин фактически не развиты, их сложно различить на длинном теле животного. Орган зрения – глаза видят плохо, их маскируют светлые участки кожи в этой зоне. В круглом рте имеются целых 2 ряда зубов, имеется также один непарный зуб районе неба. Миксины «дышат носом», при этом вода поступает в имеющееся в конце рыла отверстие – ноздрю. Органы дыхания миксин, как у всех рыб, — жабры. Зоной их расположения являются особые полости-каналы, проходящие вдоль тела животного.
По своему образу жизни миксина – животное ночное, в дневное время она зарывается в ил, прячась от других водных обитателей. При этом часть головы остается снаружи.
Охотится миксина только на тех рыб, которые больны, ослаблены (например, после нереста) или попали в снасти, сети, установленные человеком. Сам процесс нападения происходит следующим образом: миксина проедает своими острыми зубами стенку тела рыбы, после чего входит в тело, потребляя сначала внутренние органы, а потом и мышечную массу. Если несчастная жертва еще способна сопротивляться, то миксина проходит в жабры и заполняет их слизью, обильно выделяемой ее железами. В итоге рыба погибает от удушья, оставляя охотнице возможность поедать ее тело.
Нередко миксины охотятся на одну жертву совместно, однажды был зафиксирован случай, когда на одну треску напали 123 миксины. Самые любимые жертвы этих животных: осетр, треска, сельдь и скумбрия. Однако если предоставляется возможность, миксина не упустит случая полакомиться и другими рыбами.

Как правило, при достижении 25-28 сантиметров длины миксина уже является половозрелой. Миксины откладывают яйца, предварительно уходя на большие глубины. Обычно у одной самки получается кладка из 12-30 яиц, имеющих роговую оболочку для защиты. Процесс оплодотворения яиц наружный, при этом и самка, и самец продолжают нормальную жизнь после нереста. Миксины после выхода из яиц вскоре становятся полной копией родительских особей.

Поскольку внешний вид миксин и их образ питания отвратителен, никакого коммерческого промысла на нее не ведется. Более того, в местах донного травления рыба-ведьма рассматривается как экономический вредитель. В настоящее время всю пойманную миксину используют в кожевном производстве, изготавливая из нее известную «кожу угря». Особенно выделяется западное побережье Северной Америки, где уже ведется коммерческий промысел миксин.
В некоторых азиатских странах миксин используют в пищу. В Японии, Тайване и особенно в Южной Корее из нее готовят жареные блюда.
Современные исследования слизи показали, что она имеет уникальную химическую структуру. Ее состав таков, что потенциально она может заменить самые сильные средства остановки кровотечений.

Большую часть своей жизни миксины проводят на дне, предпочитая глиняную, илистую или песчаную породу. Животное копается вниз головой, приняв вертикальное положение. В донном иле они находят червей и падаль, составляющих их основной рацион питания. В тела мертвой рыбы миксины попадают через рот или жабры. Проникнув в тело, животное соскребает с костей разлагающуюся плоть. Так же не побоится напасть и на обессиленную больную рыбу, забираясь в ее внутренности таким же образом, как и в мертвое тело.
Миксину привлекает любая доступная добыча, поэтому нередко она набрасывается и на здоровую рыбу, которая не может двигаться. Рыбаки, использующие сети, знают, что в местах, облюбованных миксинами, невозможно получить хоть какой либо улов, поскольку животное атакует при любой возможности. Учитывая, что стая рыб-ведьм может содержать до десяти тысяч особей, проще сменить место рыбалки, чем надеяться, что миксины оставят добычу в покое.

ДВОЯКОДЫШАЩИЕ РЫБЫ (DIPNOI)
 Общая характеристика двоякодышащих. Жаберные области прикрыты жаберными крышками. В хрящевом скелете развиваются покровные кости (в области черепа). Хвост дифицеркальный (см. ниже). В кишечнике имеется спиральный клапан. Артериальный конус в виде извитой спиралью трубки. Плавательный пузырь отсутствует. Помимо жаберного, существует легочное дыхание. Этим признаком Dipnoi резко отличаются от остальных рыб.
Общая характеристика двоякодышащих. Жаберные области прикрыты жаберными крышками. В хрящевом скелете развиваются покровные кости (в области черепа). Хвост дифицеркальный (см. ниже). В кишечнике имеется спиральный клапан. Артериальный конус в виде извитой спиралью трубки. Плавательный пузырь отсутствует. Помимо жаберного, существует легочное дыхание. Этим признаком Dipnoi резко отличаются от остальных рыб.
Систематика. К этому подклассу принадлежат два отряда двоякодышащих рыб: 1) однолегочные и 2) двулегочные.
К первому отряду (Monopneumones) относится австралийский чешуйчатник, или цератод (Neoceratodus forsteri), распространенный в пресных водах Квинсленда (рис , А).
Цератод—самая крупная из современных двоякодышащих рыб, достигающая длины от 1 до 2 м.
Общее строение цератода. Вальковатое, сжатое с боков тело цератода заканчивается дифицеркальным хвостовым плавником, который разделяется позвоночным столбом на две почти равные половины: верхнюю и нижнюю.
Кожа одета крупной круглой (циклоидной) чешуей (без зазубренного заднего края).
Рот помещается на нижней стороне головы у переднего конца рыла; наружные носовые отверстия прикрыты верхней губой; пара внутренних отверстий (х о а н) открывается в переднюю часть ротовой полости. Наличие внутренних носовых отверстий стоит в связи с двояким дыханием (легочным и жаберным).
Строение парных конечностей замечательно: каждая конечность имеет вид заостряющегося к концу ласта.
Рис. Череп цератода сверху (левый рисунок) и снизу (правый рисунок).
1—хрящевая часть квадратной кости, с которой сочленяется нижняя челюсть; 2, 3, 4—покровные кости крыши черепа; 5—ноздри; 6—глазница; 7—praeoperculum; 8— II ребро; 9— I ребро; 10—сошниковая пластинка; 11—зубы; 12—palatopterygoideum; 13—парасфеноид; 14—interoperculum.
Скелет
Позвоночник представлен постоянной хордой совершенно не разделенной на отдельные позвонки. Сегментация выражена здесь лишь наличием хрящевых верхних отростков и хрящевых ребер.
Череп (рис.) имеет широкое основание (платибазальный тип) и состоит почти сплошь из хряща. В затылочной области отмечаются два небольших окостенения; сверху череп прикрыт несколькими поверхностными костями; снизу имеется одна большая кость, соответствующая парасфеноиду костистых рыб (рис. , 13). Небноквадратный хрящ пристает к черепу (аутостилическое соединение). Боковые части черепа с каждой стороны прикрыты височными костями (squamosum = pteroti cum; рис. 2 , 5). Жаберная крышка представлена двумя костями. Жаберные лучина хрящевых жаберных дужках отсутствуют. Плечевой пояс (рис. 2 ) состоит из толстого хряща, который облечен парой покровных костей. Скелет парных плавников слагается из основной оси, состоящей из ряда хрящиков, и хрящевых лучей, которые с каждой стороны поддерживают лопасти плавника (рис. 2 , 13). Такое строение конечности называется бисериальным. Гегенбаур полагает, что наиболее простым типом структуры конечностей надо считать скелетную ось, несущую два ряда лучей. Такую конечность этот автор называет архиптеригиумом и от нее производит конечности наземных позвоночных. По типу архиптеригиума построены парные плавники цератода.

Рис. 2 . Скелет цератода сбоку.
1,2, 3—покровные кости крыши черепа; 4—задняя хрящевая часть черепа; 5—pterotjcum (squamosum); 6—operculum; 7—suborbitale; 8—глазница; 9—плечевой пояс; 10—проксимальный хрящ грудного плавника; 11—грудной плавник; 12—тазовый пояс; 13—брюшной плавник; 14—осевой скелет; 15—хвостовой плавник.
И. И. Шмальгаузен (1915) допускает, что подобный активно гибкий с редуцированным кожным скелетом плавник цератода развился вследствие медленного передвижения и отчасти плавания в сильно заросших пресных водах.
Органы пищеварения двоякодышащих рыб
Из характерных особенностей чешуйчатника обращают на себя особое внимание его зубы. Каждый зуб представляет собой пластинку, выпуклый край которой обращен внутрь; зуб несет 6—7 острых вершин, направленных вперед. Таких зубов две пары: одна—на крыше ротовой полости, другая—на нижней челюсти. Едва ли может быть сомнение, что подобные сложные зубы произошли вследствие слияния отдельных простых конических зубов (рис., 11).
Во всю длину кишки тянется спи-ральный клапан, похожий на клапан, имеющийся у поперечноротых рыб.
Дыхание двоякодышащих рыб
Помимо жабер, у неоцератода имеется одиночное легкое, внутри разделенное на ряд камер с ячеистыми стенками. Легкое расположено на спинной стороне тела, но сообщается с пищеводом посредством канала, открывающегося на брюшной части пищевода.
Легкие неоцератода (и других двоякодышащих) как по своему положению, так и по структуре близки к плавательному пузырю высших рыб. У многих высших рыб внутренние стенки плавательного пузыря гладкие, а у двоякодышащих — ячеистые. Однако в отношении этого признака известны многочисленные переходы. Так, например, плавательный пузырь костных ганоидов (Lepidosteus, Amiа,) имеет ячеистые внутренние стенки. Повидимому, можно определенно считать, что легкие Dipnoi и плавательный пузырь высших рыб—органы гомологические.
К легкому подходят легочные артерии, а от него идут легочные вены; таким образом, оно исполняет дыхательную функцию, аналогичную лаковой у наземных позвоночных.
Кровообращение
С двойным дыханием цератода связаны характерные особенности его кровообращения. В структуре сердца обращает на себя внимание наличие на брюшной стенке предсердия перегородки, не сполна разделяющей полость предсердия на правую и левую половину. Эта перегородка вдается в венозный синус и делит его отверстие, направленное в полость предсердия, на две части. В отверстии, соединяющем предсердие с желудочком, нет клапанов, но перегородка между предсердием свисает в полость желудочка и частично прикрепляется к его стенкам. Все это сложное строение обусловливает особенности функции сердца: при сокращении предсердия и желудочка неполная перегородка прижимается к стенкам и на мгновение изолирует правые половины как предсердия, так и желудочка. Для отделения кровяного тока правой и левой половин сердца служит также своеобразное строение артериального конуса. Он спирально закручен и несет восемь поперечных клапанов, при помощи которых формируется продольная перегородка в артериальном конусе. Она отделяет левый брюшной проток конуса, по которому проходит артериальная кровь, от правого спинного, по которому течет венозная кровь.
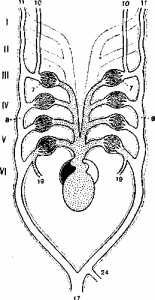
Ознакомившись со структурой сердца, легко понять последовательность в механизме кровообращения. Из легочной вены в левую часть предсердия и желудочка поступает артериальная кровь, идущая в брюшной отдел артериального конуса. От конуса берут начало четыре пары жаберных сосудов (рис 3 ). Две передние пары начинаются от брюшной стороны конуса, и потому принимают чистую артериальную кровь. От этих дуг отдходят сонные артерии, подающие чистую артериальную кровь к голове (рис. 3 , 10, 11). Две задние пары жаберных сосудов связаны со спинной частью конуса и несут венозную кровь: от задней пиры ответвляются легочные артерии, подающие венозную кровь для окисления в легкие.
Рис. 3 . Схема артериальных дуг цератода с брюшной стороны.
I, II, III, IV, V, VI—артериальные дуги; 7—жабры; 8—выносящая артерия; 10— внутренняя сонная артерия; 11 — наружная сонная артерия; 17—спинная аорта; 19—легочная артерия; 24—внутренностная артерия.
В правую половину сердца (в правый отдел венозного синуса, предсердия, а затем в желудочек) поступает вся венозная кровь, которая через протоки Кювье и через нижнюю полую вену (см. дальше) попадает в сердце.
Эта венозная кровь направляется в правый спинной венозный проток, в конус аорты. Далее венозная кровь поступает в жабры, а также в легочную артерию. Тело цератода, его внутренние органы (кроме головного отдела) получают кровь, окисленную в жабрах; в головной отдел, как было указано выше, поступает кровь, получившая более энергичное окисление в легких. Несмотря на то, что предсердие и желудочек вполне разделены на правую и левую половины, благодаря ряду описанных приспособлений, достигается изоляция чистого артериального потока крови к голове (через передние пары сосудов, отходящих от артериального конуса, и через сонные артерии).
В дополнение к сделанному очерку укажем, что в венозной системе характерно появление нижней полой вены, впадающей в венозный синус. Этот сосуд отсутствует у других рыб. Кроме того, развивается особая брюшная вена, также подходящая к венозному синусу. Брюшной вены нет у других рыб, но она хорошо развита у амфибий.
Нервная система
Для центральной нервной системы характерно сильное развитие переднего мозга; средний мозг относительно невелик, мозжечок довольно мал.
Мочеполовые органы
Почки представляют первичную почку (mesonephros); три пары канальцев предпочки функционируют только у зародыша. Мочеточники впадают в клоаку. У самок имеются парные яйцеводы в виде двух длинных извилистых трубок, открывающихся своими передними конусами (воронками) в полости тела недалеко от сердца. Нижние концы яйцеводов, или мюллеровских каналов, соединяются на особом сосочке, который открывается непарным отверстием в клоаку.
У самца имеются длинные большие семенники. У неоцератода многочисленные семявыносящие канальцы ведут через первичную почку в вольфов проток, открывающийся в клоаку. Отметим, что у самцов хорошо развиты яйцеводы (мюллеров ы протоки).
У остальных двоякодышащих имеются некоторые отличия в структуре мужских половых органов по сравнению с описанными у неоцератода. Так, у Lepido-siren семявыносящие канальцы (по 5—6 с каждой стороны) проходят только через задние почечные канальцы в общий вольфов проток. У Protopterus один задний каналец, имеющийся в наличии, вполне обособился от почки и приобрел характер самостоятельного выводного пути.
Экология. Цератод довольно обыкновенен в болотистых медленно текущих реках. Это малоподвижная вялая рыба, легко попадающаяся преследующему ее человеку. По временам цератод поднимается на поверхность, чтобы набрать воздух в легкие. Воздух втягивается с характерным звуком, напоминающим стон. Этот звук хорошо слышен тихой ночью, особенно если быть в это время на воде в лодке. Легочное дыхание является целесообразным приспособлением в период засухи, когда водоем превращается в болото: в то время много других рыб погибает, а чешуйчатник чувствует себя, повидимому, очень хорошо: в это время легочное дыхание выручает рыбу.
Необходимо отметить, что преобладающий способ дыхания у описываемого вида жаберный; в этом отношении он ближе к прочим рыбам, чем другие представители двоякодышащих. Весь год он живет в воде. Извлеченный из своей естественной среды на воздух цератод быстро погибает.
Пища состоит из мелкой животной добычи—рачков, червей, моллюсков.
Нерест с апреля по ноябрь. Яйца, окруженные студенистыми оболочками, откладываются между водными растениями.
Личинка цератода лишена наружных жабер. Интересно, что зубы не сливаются в характерные пластинки, а состоят из отдельных острых зубчиков.
3.ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ПРЕСМЫКАЮЩИХСЯ
В каменноугольном периоде широкого распространения достигли стегоцефалы. Влажный, теплый и ровный климат того времени благоприятствовал этим земноводным, обладавшим еще недостаточно совершенным легочным дыханием.
Однако в конце каменноугольного периода произошли большие поднятия земной коры, и вместе с тем на значительном пространстве суши установился сухой, местами жаркий климат. В то же время годичные кольца на стволах ископаемых деревьев того времени свидетельствуют о холодных зимах. Начали образовываться обширные пустыни. Каменноугольные леса со своими болотами и озерами постепенно сокращались. На смену пышной болотной флоре появилась хвойная и саговниковая растительность.
Новые условия среды складывались неблагоприятно для стегоцефалов. Некоторые из них, угнетаемые засушливым климатом, постепенно вымирали, другие, сохранив связь с пересыхающими водоемами, дали начало современным земноводным, третьи, вполне отошедшие от водной среды, дали начало новой ветви позвоночных—пресмыкающимся.
Пресмыкающиеся приобрели в первую очередь два чрезвычайно важных отличия от стегоцефалов. Во-первых—плотный роговой покров, предохраняющий их тело от чрезмерной потери влаги. Во-вторых—способность размножаться на суше путем откладывания яиц, получивших плотную оболочку и больший запас питательного желтка, а также особую систему зародышевых оболочек, образующуюся в процессе развития зародыша. Кроме того, все системы органов, в частности головной мозг, получили более или менее существенные прогрессивные изменения. Все это подняло пресмыкающихся на более высокую ступень организации по сравнению с земноводными.
Будучи значительно более приспособленными к условиям сухого климата и подлинно наземного существования, пресмыкающиеся уже к началу мезозоя в значительной степени вытеснили стегоцефалов. Не встречая конкурентов, они расселились повсюду, приспособляясь к самым различным условиям наземной среды. Некоторые вторично возвратились к жизни в воде. В течение всего мезозоя пресмыкающиеся занимали господствующее положение на земном шаре, достигнув, особенно в триасовом периоде, чрезвычайного обилия и многообразия форм. Мезозойская эра получила даже название века рептилий.
Древнейшими пресмыкающимися, известными уже из верхних каменноугольных отложений, были котилозавры (Cotylosanria). Они имели массивное туловище и толстые пятипалые ноги. Некоторые их них были невелики, другие достигали нескольких метров в длину. Череп котилозавров был покрыт сплошным панцирем из покровных костей с отверстиями лишь для ноздрей, глаз и теменного органа. Как по строению черепа, так и по ряду других признаков эти древние пресмыкающиеся чрезвычайно близки к стегоцефалам, которые несомненно были их прямыми предками. В то же время котилозавры обладали многими признаками настоящих рептилий. В частности, они имели один мыщелок для сочленения черепа с позвоночником. Котилозавры получили значительное развитие в пермском периоде. Среди них сеймурия (Samurai), достигавшая не более половины метра длины, считается примитивнейшей из всех известных пресмыкающихся. В триасе котилозавры вымерли.
Котилозавры дали начало всем остальным пресмыкающимся. В эволюции более поздних групп происходило значительное облегчение скелета при сохранении его прочности. В частности, это в значительной степени имело место и в черепе, костный панцырь которого подвергся частичной редукции вследствие образования височных отверстий. Редукция черепного панцыря, как уже было сказано в предыдущей главе, происходила различно. У пресмыкающихся одной группы, называемых синапсидами (Synapsida), образовалось единственное боковое височное отверстие. У рептилий другой группы (Diapsida), одновременно образовались два височных отверстия— верхнее и боковое и соответственно верхняя и нижняя височные дуги.

Сеймурия.
Краткий обзор мезозойских пресмыкающихся мы начнем с тех, которые имели лишь одну височную дугу.
Очень древнюю группу рептилий составляют черепахи (Testudines), происходящие, павидимому, непосредственно от котилозавров. Первые черепахи известны уже из пермских отложений. Это были ящерицеобразные рептилии с короткими и широкими ребрами, образующими подобие спинного щита под кожей. Брюшной щит у них еще не был развит, а челюсти были вооружены зубами. Из триаса известны уже вполне типичные черепахи. В юрское и меловое время жили многочисленные скрытошейные и бокошейные черепахи, дошедшие без существенных изменений до наших дней. Древнейшие черепахи были исключительно наземными животными, и лишь гораздо позднее некоторые из них приспособились к водному образу жизни.
Чрезвычайно своеобразную группу мезозойских пресмыкающихся составляли ихтиозавры (Ichthyosauria), Приспособившиеся исключительно к водному образу жизни. Они имели веретенообразное тело, сильно вытянутое рыло, хвост с большим двулопастным плавником и конечности в виде коротких ластов. Кожа ихтиозавров утратила роговой покров. На спине имелся кожный плавник. Позвонки были двояковогнутые, а кости таза недоразвиты. По внешнему облику ихтиозавры были похожи на дельфинов. Длина их тела колебалась от 1 до 13 метров. Ихтиозавры появились в триасе и вымерли в конце мелового периода. Они населяли моря, где питались главным образом рыбами и моллюсками. Ихтиозавры были живородящими. Происхождение этой группы пресмыкающихся до сих пор остается неясным.
Другой группой пресмыкающихся, также приспособившихся к жизни в море, были плезиозавры (Plesiosauria). В отличие от ихтиозавров, органом движения которых служил мощный хвост, плезиозавры плавали при помощи конечностей, превращенных в огромные ласты. Хвост, наоборот, был развит слабо. Более древние плезиозавры обладали ящерицеобразным сложением, позднейшие имели короткое бочонкообразное туловище, очень длинную шею и маленькую голову. Кожа у них была голая, зубы сидели в отдельных ячейках. Среди них были и мелкие формы в полметра длины и тринадцатиметровые гиганты. Плезиозавры жили одновременно с ихтиозаврами. В отличие от последних они держались, по-видимому, в прибрежной полосе моря, и, подобно тюленям, могли выходить на берег.
Крупную группу пресмыкающихся, отделившихся, вероятно, от древних котилозавров уже в верхнекаменноугольное время, составляли зверозубые (Theriodontia). Представители этой группы совмещали примитивные черты организации с рядом признаков, которые в дальнейшем становятся характерными для млекопитающих. Так, они имели двояковогнутые позвонки и зубы, сидевшие в отдельных ячейках и дифференцированные на резцы, клыки и коренные. Благодаря редукции средней части тройного затылочного бугра они приобрели двойной затылочной мысок. Многие обладали вторичным костным нёбом. И не только эти, но и ряд других особенностей зверозубых не оставляют сомнений в том, что именно они были предками млекопитающих. Развившаяся у более поздних форм способность к передвижению на высоких ногах придала и внешнему их облику удивительное сходство со зверями.
В пермском и триасовом периодах зверозубые достигли большого разнообразия. Среди них были и хищные и растительноядные формы. Некоторые зверозубые были величиной с крысу, другие достигали больших - размеров. Среди них иностранцевия—хищник с мощными клыками из пермских отложений на Северной Двине—достигала трех метров длины. Циногнат из триасовых отложений южной Африки имел до двух метров в длину.
В остальных группах мезозойских пресмыкающихся в результате частичной редукции черепного панцыря образовались две височные дуги.
Наиболее примитивными рептилиями среди диапсид считаются первоящеры (Rhynchocephalia), известные с триаса. Замечательно, что единственный; современный представитель этой группы—новозеландская гаттерия сравнительно очень мало отличается от своих древних предков.
Вероятно от общего предка с первоящерами произошли псевдозухии (Pseudosuchia), давшие в свою очередь начало крокодилам, птерозаврам, динозаврам и птицам. Это были небольшие ящерицеобразные пресмыкающиеся с зубами, сидевшими в глубоких ячейках, с неподвижной квадратной костью, но лишенные вторичного костного нёба. Псевдозухии жили в триасовом периоде, населяя главным образом Европу.
Еще в триасе от псевдозухий обособились близкие к ним древние крокодилы, но настоящие крокодилы (Crocodilia) появились только в начале юры. Вторичное костное нёбо этих пресмыкающихся претерпело длительную эволюцию. У ранних форм оно слагалось отростками челюстных и нёбных костей, у позднейших в образовании его приняли участие также отростки крыловидных костей. Вместе с тем происходило постепенное отодвигание внутренних носовых отверстий вглубь ротовой полости. Современные крокодилы мало отличаются ют своих верхнемеловых родичей.

Ихтиозавр.
Циногнат.
Чрезвычайно своеобразную группу летающих рептилий, ответвившуюся также от псевдозухий, представляли птерозавры (Pterosauria), известные из юрских и меловых отложений. Они имели крылья, образованные складкой кожи, натянутой между боками тела, и чрезвычайно удлиненным четвертым пальцем передних конечностей. Одни птерозавры — рамфоринхи — обладали узкими, длинными крыльями и очень длинным хвостом с ромбической пластинкой на конце. У других—птеродактилей—крылья были более широкие, а хвост совершенно редуцирован. Первые летали скользящим, планирующим полетом, причем хвост служил рулем. Вторые летали тяжелым гребным полетом. В связи с приспособлением к полету птерозавры приобрели ряд общих с птицами признаков, но эти признаки не были гомологичны и развились в обеих группах независимо. Широкая грудина птерозавров имела киль для прикрепления крупных грудных мышц. Кости имели воздушные полости. Вытянутые в клюв челюсти у более поздних форм были лишены зубов. Величина птерозавров была очень различна—от нескольких сантиметров до 8 метров в размахе крыльев.
Третьей чрезвычайно обширной и разнообразной группой пресмыкающихся, ответвившихся от псевдозухий, были динозавры (Dinosauria), жившие с триаса до конца мелового периода. Древнейшие представители этой группы передвигались на задних ногах, получивших преимущественное развитие. Спинной мозг динозавров имел в области крестца мощное утолщение, по объему превосходившее иногда в двадцать раз объем головного мозга, который отличался вообще очень небольшой величиной. Уже очень рано динозавры разделились на две ветви, развивавшиеся в дальнейшем параллельно. По различию в строении тазового пояса эти ветви получили название ящеротазых и птицетазых.
Древнейшие ящеротазые (Saurischia), известные из триасовых отложений, были небольшими бегающими и скачущими на задних ногах хищниками. Их скелет был очень легок, так как кости имели воздушные полости. Острые зубы были сжаты с боков и зазубрены по краю. Укороченными передними конечностями они пользовались, вероятно, лишь для схватывания добычи. Мощные задние ноги обладали длинной плюсной. Первый палец этих конечностей противопоставлялся остальным, а пятый был недоразвит. Более поздние хищные ящеротазые обладали значительно большими размерами, а позднейшие были огромными двуногими пресмыкающимися до 10 метров в длину.
В юрском периоде появились гигантские растительноядные ящеротазые, вторично вернувшиеся к хождению на четырех ногах, получивших более равномерное развитие. Они имели маленькую голову, относительно короткое туловище и очень длинные шею и хвост. Часто сменявшиеся зубы, расположенные в передней части челюстей, были приспособлены для растительной пищи. Это были не только самые гигантские из всех известных пресмыкающихся, но и вообще самые крупные наземные позвоночные, когда-либо существовавшие на земле. Среди них бронтозавр достигал 18 метров, а диплодок до 24 метров в длину.
Птицетазые (Ornithischia), распространенные с верхнего триаса до конца мелового периода, были представлены не меньшим обилием многообразных форм. Древнейшие из них также передвигались только на задних ногах, а более поздние—на четырех. Это были исключительно растительноядные рептилии. Ходивший на задних ногах игуанодон достигал не менее 5 метров высоты и был лишен кожного панцыря. Большинство других форм, наоборот, отличалось хорошо развитым панцырем, часто снабженным различными выростами, шипами, рогами и т. п. Стегозавр имел на спине двойной гребень из огромных треугольных костных пластинок. Похожий на носорога трицератопс достигал 13 метров в длину и обладал рогом на конце морды и парой рогов над глазами.
Динозавры, обнаруживая самые разнообразные приспособления и обитая в самых различных природных условиях, были распространены по всему земному шару. Достигнув наибольшего развития в мелу, в конце этого периода они вымерли совершенно.

Птерозавры.
К диапсидам примыкает группа чешуйчатых рептилий (Squamata), занимающая, однако, своеобразное положение. Древнейший представитель этой группы—Araeoscelis, известный из пермских отложений, представлял собой небольшое ящерицеобразное пресмыкающееся стройного сложения, с длинными ногами. Квадратная кость у него была подвижно сочленена с черепом. Постепенное увеличение подвижности сочленения квадратной и крыловидной костей составляло характерную особенность в эволюции чешуйчатых рептилии. Ископаемые остатки этой группы крайне скудны, но несомненно она имеет очень древнее, хотя все еще неясное происхождение. Самого Araeoscelis некоторые сближают с древнейшими первоящерами. Настоящие чешуйчатые появились в триасе. Исходными формами были ящерицы, от которых в мелу обособились змеи. В триасе и мелу чешуйчатые дали ряд боковых ветвей, приспособившихся к водной среде. Среди них мозазавры (Mosasauria) были огромными морскими пресмыкающимися с змеевидным телом и превращенными в плавника конечностями. Некоторые мозазавры достигали 15 метров в длину.
Если огромная по времени мезозойская эра заслуженно получила название века рептилий, то не менее заслуженно конец этой эры называют эпохой великого вымирания. Огромное большинство пресмыкающихся вымерло за сравнительно короткий срок в конце мелового периода, и в кенозойскую эру перешло лишь незначительное число групп этого класса, долгое время господствовавшего на суше. Причины вымирания мезозойских рептилий до сих пор не вполне ясны. Наиболее вероятным считается следующее предположение. В длительном процессе борьбы за существование пресмыкающиеся достигли узкой специализации в приспособлении к определенным условиям среды. В конце мезозоя в связи со значительными изменениями ландшафта и климата они оказались в иных условиях, в которых их узкая приспособленность утратила свою целесообразность. В то же время появились птицы и млекопитающие в качестве новых конкурентов на суше. Сложившиеся таким образом новые условия борьбы за существование привели к вымиранию рептилий и к началу расцвета более высоко организованных позвоночных
Характеристика отрядов птиц: гагарообразные, поганкообразные, буревестникообразные.
Дата добавления: 2019-01-14; просмотров: 628; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!