Организация, или «индивидуация», зачатков
Типичная последовательность расчленения зачатка нервной системы определяется, по-видимому, также некоторым взаимодействием. Для нормальной организации зачатка нервной системы требуется, прежде всего, известная длительность взаимодействия эктодермы с хордомезодермой. В эктодерме сначала индуцируется лишь способность к развитию нервных тканей, а затем постепенно определяется способность к образованию нервной трубки и, наконец, детерминируются определенные отделы головного и спинного мозга. Иногда различают две формы индукции — эвокацию, т. е. влияние, вызывающее образование зачатка, и индивидуацию, т. е. влияние, вызывающее региональные различия и общую организацию зачатка. На конкретном материале трудно разграничить эти процессы, и вряд ли между ними имеются принципиальные различия. Весь процесс детерминации идет под влиянием контакта с тем же индуктором.
На стадии гаструлы при повороте эктодермы будущей нервной пластинки на 180° она образует все же под влиянием индуктора нормальную последовательность отделов мозга. На стадиях ней-руляции (закладка нервной пластинки) при таком же опыте образуется обратная последовательность отделов. Последовательность расчленения устанавливается, следовательно, одновременно с окончательной детерминацией материала нервной пластинки, очевидно, под влиянием того же индуктора. В таком случае нужно ожидать, что индуцирующие свойства различных отделов хор-Домезодермы различны. Это предположение показано экспериментально как в опытах с гомопластическими пересадками (рис. 12), так и в опытах с гетеропластической эксплантацией разных отделов хордомезодермы (рис. 13). В последнем случае для эксперимента были взяты части хордомезодермы одного вида тритонов и
|
|
|
256 Регуляция формообразования в индивидуальном развитии
большие куски индифферентной еще эктодермы ранней гаструлы другого вида, отличающегося по размерам и пигментации клеток. Куски хордомезодермы завертывались в эктодерму, и такая искусственная система культивировалась в солевом растворе. В результате развивались более или менее сложные образования с самыми различными органами, происхождение которых можно было точно проверить по характеру клеток. При эксплантации в такой системе материала верхней губы бластопора или образовавшейся из нее передней части хордомезодермы этот материал дифференцируется в переднюю (головную) кишку, хорду, мускулатуру. В прилежащем эктодермальном покрове (другого вида)

Рис. 12. Действие головного и хвостового индукторов [Кюн, 1955]
1—3 — схемы операций; 4 — действие головного индуктора, операция по схеме 1-+3; 5 — действие хвостового индуктора, операция по схеме 2-*-3
|
|
|
развиваются части головного мозга, глаза, обонятельные ямки и иногда слуховые пузырьки, т. е. только типичные головные органы. При эксплантации кусочка задней части хордомезодермы из нее в таком же опыте развиваются хорда и мускулатура, а в прилежащей эктодерме только зачаток спинного мозга, т. е. туловищные органы. В связи с концом задней части развиваются и более или менее хорошо выраженные хвосты с продолжением хорды и мускулатуры. Следовательно, передняя и задняя части индуктора действительно различаются по своим свойствам. Дальнейшие опыты показали, что между разными отделами хордомезодермы нет качественных различий. Имеются лишь какие-то количественные различия и постепенное изменение свойств при переходе от переднего конца крыши первичной кишки к заднему. Быть может, эти различия связаны с последовательностью в развитии самого материала. Передний конец образуется из того
| 257 |
Глава II . Регуляция формообразования у земноводных
материала верхней губы бластопора, который ранее всего ввертывается внутрь и проделывает довольно длинный путь под эктодермой до района анимального полюса. Задний конец крыши образуется позже всего — в самом конце гаструляции.
|
|
|
Таким образом, мы приходим к вероятному выводу о том, что региональное расчленение зачатка нервной системы зависит, по крайней мере отчасти, от свойств самого индуктора, который также обнаруживает региональные различия. Однако в индукторе эти различия имеют количественный характер постепенного изменения от переднего конца к заднему. По-видимому, такой характер различий определяется изменением концентрации каких-то

Р^с. 13. Эксперименты с помещением испытуемого индукционного материа-л« в презумптивную эктодерму [Кюн, 1955]
2 > 4 — схемы операций; 2, 3 — результаты операций по схеме 1; 2—головной «организатор» тритона в эктодерме жерлянки; 3 — головной «организатор» жерлянки в эктодерме жерлянки; 5, 6—результаты операции по схеме 4\ 5 — туловищный Организатор» жерлянки в эктодерме тритона; 6 — то же, в разрезе; Af — задняя кищКа; Аи — глазной бокал; Ch — хорда; Л—плавник; Geh — мозг; L — хруста-Лик; Ми — мускулатура; IV — нервная трубка; Na — эпителий носовой ямки
258 Регуляция формообразования в индивидуальном развитии
веществ или изменением процессов клеточного обмена. Между тем, в зачатке нервной системы под влиянием этих количественных различий наступает разделение на качественно различные отделы центральной нервной системы (развитие которых определяется пороговым уровнем поступления индуцирующих веществ). Формообразовательное значение количественных различий в распределении известных веществ, связанное с изменением метаболизма в известных направлениях, установлено уже во многих случаях. Постепенное падение или нарастание концентрации некоторых веществ, проявлений метаболизма, индуцирующих свойств или способности к известным реакциям (компетенции) характерно для очень многих зачатков и обозначается как падение или нарастание некоторого градиента. Известный американский исследователь Чайльд пытался построить на явлениях градиента законченную общую теорию развития. Это вряд ли возможно. Однако огромное значение градиентов совершенно бесспорно. Между прочим, именно наличие градиента дает прекрасное объяснение высокой способности к регуляции всех ранних, но уже детерминированных в общем зачатков органов, так как при удалении части зачатка общий градиент не нарушается и вновь определяет развитие целого.
|
|
|
Природа индуцирующих влияний
Опытами межвидовой пересадки было доказано отсутствие видовой специфичности индуктора: индукция у тритона может быть вызвана материалом, взятым от других видов земноводных (например, лягушки) или даже от очень далеких животных, как, например, птиц и даже млекопитающих.
Более того, индукция нервной системы может быть вызвана в недетерминированной (индифферентной) эктодерме не только материалом крыши первичной кишки (хордомезодермы) или его производными — хордой и мышцами, но и кусочком нервной пластинки. Это напоминает явления ассимиляторной индукции, однако в отличие от последней индуцированная ткань не вовлекается в состав ткани самого индуктора. Было найдено, что и различные другие ткани, как, например, кусочки печени, могут вызвать образование нервной пластинки.
Это заставляло думать, что явление индукции связано с передачей каких-то широко распространенных в различных тканях химических веществ, которые могли бы сохраняться и в убитых тканях. Действительно, оказалось, что губы бластопора, растертые, убитые нагреванием, кипячением или даже действием спирта, также могут вызвать образование нервной пластинки. Раздавливали материал верхней губы бластопора и центрифугированием отделяли более активные части протоплазмы, которые также хорошо индуцировали. Наконец, извлекали индуцирующие вещест-
Глава II . Регуляция формообразования у земноводных 25$
ва химическими методами и наблюдали нейральную индукцию в эктодерме гаструлы под влиянием таких веществ. В настоящее время химическая природа индуцирующих веществ не вызывает сомнений. Многочисленными опытами показано, что различные вещества, извлеченные из разных тканей, различаются и по результатам индукции. Из печени морской свинки было извлечено > (петролейным эфиром) вещество, содержащее жирные кислоты и нуклеопротеиды, которое индуцировало в гаструле тритонов органы передней части головы (передний и промежуточный мозг, глаза, органы обоняния), а из почек той же морской свинки получен после подобной же экстракции нерастворимый остаток, который вызывает индукцию туловищных органов (спинного мозга, хорды, мускульных сегментов, почечных канальцев, хвостов) . В этом виден интересный параллелизм с влиянием переднего и заднего отделов хордомезодермы, которые, возможно, также отличаются и по составу действующих веществ (точнее по их относительной концентрации).
Что касается способов передачи этих веществ при нормальном развитии, то она осуществляется, очевидно, путем диффузии растворимых веществ или даже путем активного перемещения гранул по межклеточным мостикам. Прямые наблюдения показывают, что в плазме клеток в области бластопора происходит накопление большого количества гранул, содержащих рибонуклео-протеиды и переходящих при инвагинации в область крыши первичной кишки. Эти гранулы накапливаются между хордоме-зодермой и прилежащей эктодермой. Затем они появляются в большом числе в клетках эктодермы. Прижизненная окраска гранул крыши первичной кишки с помощью нейтрального красного показывает, что они перемещаются от индуктора к реагирующей ткани и, вероятно, непосредственно определяют ее преобразование в нервную пластинку. Интересное подтверждение этих выводов было получено в опытах с изолированными культурами. Если эктодерма гаструлы культивируется в среде (солевом растворе), в которой до этого в течение нескольких дней культивировался хордомезобласт, то в ней дифференцируются нервные и пигментные клетки без всякого организатора (пигментные клетки нормально развиваются из боковых частей нервной пластинки, дающих начало также нервным узлам). Оказывается, что солевой раствор после культивирования хордомезодермы сильно поглощает ультрафиолетовые лучи и содержит рибонуклеопротеиды.
Все эти факты, выявляющие химическую природу индуцирующих веществ и возможные пути их передачи, показывают вместе с тем и их малую специфичность. Это обычные клеточные субстанции, которые, однако, присутствуют в индукторе в весьма большой концентрации. Эти вещества могут быть заменены другими веществами с аналогичным действием, во всяком случае
260 Регуляция формообразования в индивидуальном развитии
они, по-видимому, одинаковы у самых различных животных. С другой стороны, реакция на действие индуктора строго специфична. Она не только различна в различных реагирующих тканях, но обладает и видовой спецификой, ясно обнаруживаемой в строении индуцированных органов.
Если у молодого зародыша тритона эктодерму передней части головы заменить эктодермой, взятой от зародыша лягушки, то
|
|

| Рис. 14. Образование балансера у зародыша жабы при пересадке эктодермы тритона в область рта [Уод-дингтон, 1956] |
под влиянием индуктора (передней кишки) тритона в эктодерме разовьются ротовые органы. Однако соответственно происхождению эктодермы от лягушки из нее разовьются у тритона типичные органы лягушки — присоски и роговые зубы. В другом эксперименте, при пересадке эктодермы тритона на голову зародыша жабы (рис. 14), в ней под влиянием индуктора жабы развиваются балансеры — типичные личиночные органы тритона. Специфическая видовая дифференцировка органов и гистологических структур во всех таких опытах определяется всегда не индуктором, а происхождением реагирующего материала. Индуктор не обладает видовой специфичностью, и он может быть заменен индукторами совершенно других организмов. Реагирующая ткань всегда выявляет видовые особенности согласно своему происхождению. Это показывает, что реализация формообразовательных процессов всегда контролируется наследственными свойствами материала — его нормой реакций.
В свете изложенных фактов становится ясно, что говорить о «первичном» индукторе как об «организаторе» — неправильно. Однако даже если его роль сводится лишь к активации некоторых дремлющих (латентных) возможностей или тенденций развития реагирующего клеточного материала, то его значение нельзя и недооценивать. Индуктор или эвокатор, во всяком случае: 1) решает (реализацией контакта и передачей активирующих веществ) вопрос альтернативного выбора одного из возможных путей развития; 2) фиксирует активированный путь развития и одновременно затормаживает другие пути. Кроме того, и это, может быть, самое существенное, установление временного контакта между индуктором и реактором приводит к согласованию времени, места и протяженности формообразовательных процессов во взаимодействующих частях.
В этом смысле весьма показательны результаты следующего
Глава II . Регуляция формообразования у земноводных 261
эксперимента. Эктодерма от вида тритонов с крупными яйцами пересаживалась в область крыши первичной кишки другого вида—с мелкими яйцами. Нервная пластинка, развившаяся под влиянием контакта, имела, конечно, крупноклеточное строение, общие же ее размеры соответствовали размерам малого вида. Таким образом, здесь сразу установились нормальные соотношения между органами, т. е. пропорциональное целое.
Мы рассмотрели пока только вопрос о значении первичного индуктора — верхней губы бластопора и развивающейся из нее крыши первичной кишки. В результате этой индукции в эктодерме образуется нервная пластинка, которая в свою очередь получает свойства индуктора. В эксперименте она может как мы уже упомянули, индуцировать и новую нервную пластинку. В нормальном развитии она расчленяется на отделы головного и спинного мозга, дает начало ганглионарной пластинке, а через это — ганглиям периферической нервной системы, пигментным клеткам и даже некоторым частям скелета (висцеральному скелету головы).
Различные части нервной системы обладают также индукционными свойствами. Передний край нервной пластинки индуцирует в прилежащей эктодерме образование органов обоняния. Задний мозг индуцирует в близлежащей эктодерме развитие слуховых пузырьков, т. е. органа слуха. В области промежуточного мозга за счет самой нервной пластинки развиваются зачатки глаз — глазные пузыри, которые затем превращаются в глазные бокалы. Мы уделим особое внимание именно развитию глаза, как наиболее изученному объекту экспериментальных исследований.
Саморазвитие глаза
Зачатки глаза обладают свойствами индуктора. Когда глазной бокал, растущий от зачатка мозга, касается эктодермы, на месте контакта эктодерма утолщается и превращается в зачаток хрусталика глаза (рис. 15). Если до этого сместить глазной бокал так, что он не может достигнуть эктодермы, то хрусталик не развивается. Если зачаток глаза пересадить на любое другое место под эктодерму, то на этом, чуждом для него месте развивается все же нормальный хрусталик. То же самое произойдет, если на ранней стадии развития глазного зачатка заменить головную эктодерму туловищной. Из туловищной эктодермы на месте контакта с глазным бокалом развивается типичный хрусталик. Зависимость развития хрусталика от индуцирующего влияния глазного бокала совершенно ясна (у большинства амфибий). Однако этим дело не ограничивается. В самом зачатке хрусталика наступает дифференцировка его клеток опять-таки в зависимости от инДуцирующего влияния внутренней стенки глазного бокала, т- е. зачатки сетчатки глаза. В хрусталике, имеющем вначале виц
| 262 Регуляция формообразования в индивидуальном развитии |

Рис. 15. Нормальное развитие глаза у аксолотля [Кюн, 1955] 1—4 — стадии развития
полого пузырька, утолщается внутренняя стенка, обращенная к сетчатке, и истончается наружная стенка. Клетки внутренней стенки превращаются в прозрачные волокна, образующие волокнистое ядро хрусталика, которое и служит светопреломляющим телом глаза. Клетки наружной стенки образуют тонкий прозрачный покров. Под влиянием глаза происходит также преобразование прилежащей кожи в прозрачную роговицу. Опыты пересадки доказали и здесь наличие индуцирующих влияний со стороны глаза.
Как и в опытах с первичным индуктором, все эти влияния малоспецифичны и могут быть заменены другими сходными влияниями. Образование хрусталика может быть вызвано различными тканями, экстрактами и веществами, подобными головным индукторам. Дифференцировка хрусталиковых волокон, и притом в строгой полярной ориентировке, наступает не только при контакте с ретиной, но и при контакте с чувствующим эпителием слухового пузырька, слуховым ганглием и другими нервными тканями (рис. 16). Очевидно, суть заключается в сходных или даже идентичных веществах, выделяемых сходными органами.
С другой стороны, и в этих опытах проявляется строгая видовая специфичность реагирующей ткани. При обменных пересадках между зародышами разной величины, например двумя видами тритонов — большим (Tr. cristatus) и малым (Tr. taeniatus) или аксолотлем и тритоном, наблюдаются, однако, не только сохранение специфических свойств клеточного материала, но и интересные соотношения в общих размерах. Эпидермис аксолотля образует при контакте с глазным бокалом тритона, согласно своему происхождению, чрезмерно большой хрусталик. Эпидермис малого тритона (Tr. taeniatus) образует в глазу большого тритона (Tr. cristatus) слишком малый хрусталик. Эпидермис большого тритона образует под индуцирующим влиянием глазного бо-
Глава II . Регуляция формообразования у земноводных 263
кала малого тритона слишком большой хрусталик, рост которого, однако, при дальнейшем развитии несколько задерживается. В этом видна некоторая способность к регуляции роста. Она находит свое выражение также в регуляторном росте глазного бокала и указывает на наличие обратного влияния со стороны хрусталика на глазной бокал. Так, в случае развития большого хрусталика из эпидермиса аксолотля в контакте с малым бокалом тритона последний начинает также увеличиваться в своих размерах. Согласование величины глазного бокала и хрусталика есть один из важнейших результатов их взаимовлияния. При удалении одного бластомера с половиной серого полумесяца (т. е. первичного индуктора), из второго бластомера развивается совершенно нормальная личинка половинного размера. При пересадке глазного бокала такого половинного размера в личинку нормального размера, в ней индуцируется и хрусталик половинного объема соответственно пересаженному бокалу.
|
|
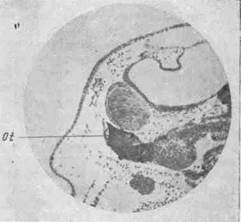
Рис. 16. Индукция хрусталико-вого ядра слуховым пузырьком 'Драгомиров, 1929]
Ot — радужина трансплантированного глаза
Рис. 17. Эксплантация зачатка глаза, завернутого в эктодерму Лопашов, I960]
1 — эксплантация зачатка глаза
без мезенхимы; развилась только сетчатка без пигментного эпителия;
2 — то же, вместе с боковой мезо-
дермой; почти весь зачаток дал сетчатку. Только в месте подхода тяжей мезенхимы развилась пигментная оболочка
|
|

264 Регуляция формообразования в индивидуальном развитии
При оплодотворении яйца, из которого предварительно было удалено ядро, получаются зародыши, клетки которых содержат лишь половину нормального числа хромосом. Такие гаплоидные клетки имеют вдвое меньшие размеры. Если эпидермис такого гаплоидного зародыша пересадить к глазному бокалу нормального диплоидного зародыша, то из него развивается хрусталик нормальной величины, состоящий из значительно большего числа маленьких гаплоидных клеток.
Величина хрусталика определяется, следовательно, не величиной клеток и не их числом, а общей нормой реагирования данного вида на данный индуктор. Если обе части реагирующей системы принадлежат одному виду, то сразу образуется пропорциональное целое. В результате индуцирующего влияния глазного бокала развивается согласованная система: глазной бокал — хрустали^ которая в значительной степени способна к регуляции своей структуры. Дело не ограничивается соотношениями в величине его частей. Происходит и взаимный контроль гистологической дифференцировки.
Глаз является относительно простой формообразовательной системой. Наружное его положение, легкость пересадок и эксплантации привели к тому, что механизм развития глаза и факторы морфологической и гистологической дифференцировки его частей оказались изученными с гораздо большей полнотой, чем какие бы то ни было другие формообразовательные процессы. Вместе с тем результаты экспериментов имеют и общее значение — они выяснили сложность взаимоотношений, определяющих саморазвитие уже детерминированной системы. Глазной пузырь детерминирован лишь в общем. В эксперименте любая его часть может дать начало ретине или пигментной оболочке. Направление развития диктуется условиями, в которые попадают эти части (рис. 17). Если в солевом растворе части глазного пузыря или бокала соединяются в сплошную массу, они развиваются только в сетчатку. Если же эти части растягиваются и располагаются тонким слоем, то они сетчатки не дают, а в присутствии мезен-химных клеток превращаются в пластинки или пузыри из пигментного эпителия. В нормальном развитии глазного пузыря его наружная стенка с самого начала несколько толще и прилегает вплотную к эктодерме покровов. Естественно, что. в сплошиов массе клеток легко происходит накопление продуктов их жизнедеятельности, а снабжение кислородом затруднено. Это определяет развитие глазного пузыря в сетчатку. Тесная связь с эктодермой и образование пограничной мембраны приводят к стягиванию растущей стенки глаза. Это способствует ее свертыванию в бокал. Одновременно в эктодерме образуется хрусталик, который выполняет полость бокала. Все эти тесные связи поддерживают наметившееся развитие в сторону сетчатки. По мере ввертывания наружной стенки пузыря внутрь она становится внутренней
Глава II . Регуляция формообразования у земноводных 265
стенкой бокала и оказывается в условиях весьма полной изоляции от внешней среды, так как вход в бокал замыкается хрусталиком. Наоборот, обращенная к мозгу стенка пузыря оказывается снаружи и растягивается по мере образования бокала и его дальнейшего роста. Получается тонкий слой клеток, свободно омываемый внешней средой. Естественно, что продукты жизнедеятельности клеток свободно удаляются и, с другой стороны, происходит свободное снабжение кислородом. Как и в солевых культурах, такое положение не допускает дифференцировки ретины и способствует образованию пигментной оболочки. Для реализации этой возможности требуется, однако (как и в культурах), наличие мезенхимных клеток, которые обволакивают глазной бокал одним слоем, образующим род очень тонкой мембраны. Позднее в этот слой мезенхимы врастают кровеносные сосуды (образуется сосудистая оболочка глаза), и лишь тогда, при полном обеспечении снабжения кислородом, пигментная оболочка глаза достигает своего полного развития. На стадиях глазного пузыря и раннего глазного бокала ткани их стенок еще не детерминированы в своих частных свойствах — они одинаково способны дать начало в соответствующих условиях как сетчатке, так и пигментной оболочке. Любой участок глазного бокала может дать начало и небольшому целому, правильно сформированному, глазу. При дифференцировке внутреннего слоя бокала в ретину происходит ряд изменений (между прочим, выталкивание имеющихся и в нем пигментных зерен), после этого клетки этого слоя оказываются уже окончательно детерминированными — они могут теперь развиваться только в сторону образования вполне дифференцированной сетчатки. Наоборот, клетки пигментной оболочки глаза у земноводных вообще не достигают уровня прочной детерминации и сохраняют свою способность дать в подходящих условиях начало как ретине, так и более сложным глазным структурам и даже целому глазу. Эта способность сохраняется у бесхвостых амфибий вплоть до метаморфоза, а у хвостатых — в течение всей жизни.
Мы дали лишь схематический обзор факторов, определяющих дифференцировку некоторых частей глаза. В действительности соотношения еще много сложнее. Приведенных данных, однако, Достаточно для ряда важнейших выводов. Клеточная дифферен-Цировка определяется различиями в обмене веществ, которые Устанавливаются при данном положении частей. Эти различия связаны с соотношением частей между собой и с окружающей их средой. В частности, отмечено значение механических факторов стягивания и растяжения, проницаемости оболочек, условий накопления или удаления продуктов собственного обмена, степени снабжения кислородом. Во всех этих случаях ярко выступает значение исторически сложившихся взаимозависимостей для нормального саморазвития данной системы.
266 Регуляция формообразования в индивидуальном paneutuu
Поясним это положение еще только одним примером. Части нервной пластинки дифференцируются при культуре в солевых растворах в зависимости от условий, в какие они попадают. Кусочки, приставшие к дну сосуда или к куску хорды, растягиваются и образуют тонкие пластинки. При контакте с мезенхимой образуются тонкостенные пузыри и трубки. В изолированных кусках и при контакте с сомитами (мускульными сегментами) образуются толстостенные участки мозга. В нормальном развитии нервной пластинки боковые части, прилежащие к сомитам, утолщаются. Средняя, прилежащая к хорде часть образует тонкую пластинку. После замыкания нервной пластинки в трубку спинной шов этой трубки формируется под влиянием мезенхимы и дает начало тонкой эпителиальной крыше. В толстых боковых массах идет, очевидно под влиянием накопления, собственных продуктов, самодифференцирование нервной ткани. Эти зависимости в общем сходны с теми, которые характерны и для развития глазного бокала. И в данном случае ясно выступает значение исторически сложившихся взаимоотношений развивающегося органа с его нормальным окружением.
Взаимосвязи оказывают формообразовательное воздействие как через механические влияния, так и, в особенности, через изменение в клеточном метаболизме. При индукции нервной пластинки это происходит, очевидно, путем непосредственной передачи рибонуклеопротеидов. В других случаях передаются, вероятно, более простые продукты обмена, или происходит накопление собственных продуктов, если для их удаления имеются препятствия. Не меньшее значение имеет, конечно, приток питательных веществ и снабжение кислородом. Во всех этих случаях небольшие, даже количественные различия приводят к изменению обмена веществ, дальнейшему нарастанию различий и, наконец, к качественным различиям в дифференцировке первоначально однородных зачатков.
Мы особо подчеркиваем, что все формообразовательные влияния имеют внутри данной системы характер сложных взаимозависимостей. Мы уже обращали внимание на то, что хордомезо-дерма определяет развитие нервной пластинки, но только обратное влияние нервной пластинки окончательно определяет развитие хорды и мезодермы в крыше первичной кишки. Различные части хордомезодермы вызывают развитие разных отделов мозга, а мускульные сегменты — образование ганглиев. С другой стороны, разные отделы мозга вызывают развитие органов чувств, которые окончательно формируются во взаимодействии с передней частью крыши первичной кишки. Глазной пузырь подрастает под эктодерму и связывается с нею. Это вызывает в эктодерме образование хрусталика, но оказывает и обратное влияние на глазной пузырь, определяя его преобразование в бокал. Внутренняя стенка бокала, изолированная хруста-
Глава II . Регуляция формообразования у земноводных 267
ликом от внешнего окружения, развивается в ретину, которая оказывает обратное влияние на хрусталик, определяя в ближайшей его стенке образование волокнистого ядра. Однако и в дальнейшем, только при взаимном влиянии ретины и хрусталика завершается их окончательная дифференцировка. В самом глазном пузыре его стенки первоначально равнозначны. Однако с образованием бокала они попадают в разное положение и характер их обмена явно расходится. В условиях сжатия клеток, накопления продуктов обмена и малого снабжения кислородом (чему способствует сначала связь с эктодермой, а затем положение хрусталика) во внутренней стенке бокала дифференцируется ретина. В условиях растяжения внешней стенки бокала, свободного удаления продуктов обмена и хорошего снабжения кислородом из нее развивается пигментная оболочка. Последняя развивается в зависимости от условий формообразования и роста внутренней стенки — ретины. Однако и пигментная оболочка оказывает свое влияние на ретину. Между нею и ретиной имеется щелевидное пространство (остаток полости глазного пузыря) с жидкостью, в которую, с одной стороны, могут удаляться продукты обмена из ретины и, с другой стороны, может подводиться кислород. И то, и другое происходит через посредство пигментной оболочки. Это определяет развитие слоя зрительных рецепторов в ретине на поверхости, обращенной к пигментной оболочке (высшие дифференцировки возникают лишь в условиях максимальной изоляции на противоположной стороне ретины, обращенной внутрь глаза; здесь развивается слой нервных клеток).
Резюмируя все сказанное, мы должны отметить, что саморазвитие известной системы определяется клеточным обменом веществ в ее частях и тканях. Характер клеточного обмена зависит прежде всего от веществ (энзимов, рибонуклеопротеидов и т. п.), имеющихся в составе цитоплазмы этих частей или тканей и полученных либо прямо от цитоплазмы яйца при его дроблении, либо синтезированных при участии активных веществ цитоплазмы. Он зависит и от продуктов обмена, накапливающихся в клетках данной ткани или поступающих в данные клетки извне — из соседних тканей. Все это — процессы, зависящие от пространственных соотношений и меняющиеся во времени. В больших массах клеток дифференцировка идет всегда полнее. С течением времени эти процессы нарастают — длительный контакт взаимодействующих частей ведет к более полной дифференцировке. И в изолированной массе однородных клеток начавшаяся уже Дифференцировка продолжает нарастать. Даже в солевых культурах происходят возрастные изменения. Последние, как мы видели, имеют большое значение, определяя компетенцию эмбриональной ткани, т. е. ее способность к определенным реакциям. Вместе с тем отметим, что нормальное развитие формообразовательных систем возможно только в условиях нормальных, исто-
268 Регуляция формообразования в индивидуальном развитии
рически сложившихся соотношений. Это, в свою очередь, говорит в пользу того, что формообразовательные взаимодействия, все средства связи, специфика активных веществ и формы реагирования сами приобрели свое значение только вместе с историческим развитием данных систем.
Во всех случаях эмбрионального развития при нормальных соотношениях частей взаимодействия внутри системы как между клетками данной ткани, так и между различными зачатками и тканями имеют характер взаимной стимуляции процессов, ведущих к прогрессивному развитию данной системы. Как мы отмечали в введении, это характерно для систем регуляции с положительной обратной связью.
Дата добавления: 2019-01-14; просмотров: 277; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!