Выпадение излишних формообразовательных реакций как кажущееся закрепление модификаций (аккомодаций)
Мыотметили в предыдущем, что характер модификационной изменчивости вырабатывается исторически (через естественный отбор) в зависимости от тех условий, в которых живет организм. Если эти условия разнообразны и изменчивы и организм не обладает средствами быстрой защиты или изоляции от резких изменений среды, то обычно модификационная изменчивость приобретает большой размах и иногда приводит к своеобразному модификационному полиморфизму (многие растения).
Если такой индивидуально весьма пластичный организм меняет свою среду (т. е. либо сама среда изменяется, либо организм переселяется в иную среду) на более постоянную, то некоторая его модификационная реакция станет почти постоянной, а другие, рассчитанные (в своем историческом развитии) на такие условия среды, которые здесь уже не встречаются, окажутся совершенно излишними. Однако мы в первой части настоящей работы уже отметили, что реакционные и корреляционные системы, определяющие развитие организма и его частей, не только вырабатываются в процессе естественного отбора, но и поддерживаются при его постоянном участии. Механизмы, потерявшие свое значение для организма, не контролируются отбором, так как их нарушения не сказываются на жизнеспособности организма в данных условиях существования. Поэтому в этих реакционных и корреляционных системах происходит беспорядочное накопление мутационных отклонений (и в первую очередь сдвигов во времени наступления известных реакций), которые приводят эти механизмы к расстройству. Распад реакционных и корреляционных систем имеет своим неизбежным следствием сначала недоразвитие, а затем и полное выпадение тех признаков и черт
|
|
|
Глава IV . Прогрессивная эволюция. Адаптациогенез 145
организации, развитие которых ими контролировалось (редукция органов). Вследствие распада установившегося внутреннего механизма развития исторически приобретенная способность к адаптивной реакции уже не может осуществиться даже при наличии того внешнего раздражителя, который раньше определял наступление этой формообразовательной реакции. Таким образом, у организма сохраняется лишь тот фенотип, который соответствует условиям развития в новой среде. Другие его исторически (в свое время) сложившиеся формы реагирования (модификации) теряются. Если у организма имеется только один ярко выраженный фенотип, то обычно говорят о наследственной структуре (неправильно, так как наследственна лишь норма реакции при данных условиях), и в данном случае, при утере других (якобы ненаследственных) фенотипов, ламаркисты говорят о наследственном «фиксировании» модификаций, о превращении их в мутации и т. п.
|
|
|
В. Кирпичников [1935] выдвигает для объяснения тех же явлений утраты излишних приспособлений гипотезу косвенного отбopa, действующего при эволюции всего организма через систему корреляционных зависимостей также разрушающим образом на механизмы, утратившие свое значение. Эти совершенно правильные соображения должны быть приняты во внимание. Результат получается тот же самый. Мне, однако, кажется, что распад вследствие отсутствия отбора как фактора, поддерживающего данный реакционный механизм, должен происходить быстрее — даже без других изменений, приобретаемых в силу положительного отбора (а без этого не может сказаться его косвенное разрушающее влияние), так как мутации отрицательные, нарушающие развитие, вообще встречаются неизмеримо чаще, чем положительные.
На каком-либо более конкретном примере нетрудно показать полную ошибочность мнения ламаркистов о фиксировании модификации. Возьмем сначала пример растения с довольно широкой нормой реакций, т. е. обладающего значительной модификацион-ной изменчивостью (объясняющейся разнообразием условий, в которых оно живет, например, при широком распространении семян ветром). Возьмем, например, обыкновенный одуванчик, который растет где угодно, в том числе и в горных местностях (рис. 17). В долинах он имеет всем нам знакомый вид, а высоко в горах Дает карликовую форму, напоминая по виду многие другие альпийские растения [Боынье, 1890]. Эта горная модификация одуванчика имеет адаптивный характер — низкорослость (близость к нагретой почве) предохраняет его от вымерзания при ночном охлаждении воздуха, а глубокие корни способствуют использованию небольших количеств воды. Если бы это растение не обладало приспособлением для широкого распространения семян при Помощи ветра, то оно могло бы, так сказать, прочно осесть в го-
|
|
|
146 Организм как целое в индивиду ильном и историческом развитии
pax. В таком случае реакционная система развития высокорослой долинной модификации оказалась бы излишней и в силу беспорядочного накопления мутаций (при отсутствии естественного отбора по долинной модификации) подверглась бы распаду. Дан-пая реакционная система будет утрачена, и нормальная долинная модификация не разовьется при переносе такого горного

Рис. 17. Долинная (Р) и горная (М) модификации одуванчика ( Taraxacum officinale ) при одинаковом уменьшении (1/5) [Циммерманн, 1938]
|
|
|
М1 — горная модификация в натуральную величину
Глава IV . Прогрессивная эволюция. Адаптациогенез 147
растения в долину (это не значит, что оно вообще не будет реагировать на такое изменение среды, но реакция эта потеряет свое прежнее адаптивное оформление). Получается впечатление, как будто теперь горное модификационное изменение адаптивного характера стало почти прочным наследственным изменением, т< е. как будто модификация закрепилась и превратилась в длительную модификацию или даже в мутацию. Мы и в самом деле получили новую наследственную форму, приближающуюся к горной, но это не есть результат «закрепления» торной модификации. Горная модификация имела и раньше наследственную базу для своего развития, которая полностью реализовалась при условиях горного климата. Новая форма отличается от старой лишь утратой своей прежней долинной модификации, которая тоже имела наследственную базу для своего развития; но теперь эта база разрушена благодаря бесконтрольному накоплению мутаций. Следовательно, и генотип изменился. Конечно, мы не можем ожидать, чтобы в долине выросло типичное горное растение, но по отсутствию типичных черт долинной формы оно будет к нему приближаться. С другой стороны, допустим, что модификационно-полиморфное растение, подобное одуванчику, осело прочно в долине (что возможно лишь при утрате способности к легкому переносу семян). В этом случае оно утеряет свою горную модификацию, так как эта форма реагирования не контролируется более естественным отбором, и данная реакционная система подвергнется распаду. Получится наследственно новая долинная форма, производящая впечатление наследственного закрепления модификационных признаков. В самом деле, и здесь лишь выпадение известного наследственного свойства отвечать определенной формообразовательной реакцией на условия горного климата. Конечно, нельзя ожидать, чтобы это долинное растение при переносе в горы не изменилось и полностью сохраняло бы свой долинный вид. Растение будет реагировать на новые условия, однако вряд ли эта реакция будет похожа на прежнюю и скорее всего растение в горах просто погибнет. Теперь обе наши новые формы — долинная и горная — обладают наследственными различиями. При скрещивании их друг с другом в потомстве получится, вероятно, и картина менделевского расщепления.
На основании наших теоретических соображений относительно эволюции доминантности мы можем даже предсказать, что при скрещивании горной формы с долинной и взращивании гибридов в долине долинная форма будет доминировать, так как механизм развития горной формы не сможет реализоваться при данных условиях развития. Наоборот, при взращивании того же гибрида в горах, доминировать, очевидно, будет горная форма, так как Долинная реакция, механизм которой унаследован от одного из родителей, не сможет реализоваться в данных условиях. Одна четвертая часть особей, содержащая в чистом виде механизм
148 Организм как целое в индивидуальном и историческом развитии
долинной реакции, даст, конечно, не типичную долинную форму но и не горную, а какую-либо мало приспособленную модификацию. Сказанное можно пояснить схемой (рис. 18).
Довольно много конкретных примеров такой постепенной утери прежних реакционных механизмов приведено у Вайдеи-райха [ Fr. Weidenreich, 1921]: альпийские растения, дающие полную долинную реакцию, частично или полностью ее потерявшие

Рис. 18. Схема дивергентной эволюции через потерю частных реакций при переселении в различные среды как кажущееся закрепление модификаций
растения, перешедшие от гигрофилии к ксерофилии, но не утерявшие способности давать гигрофильную реакцию (например, листья вместо колючек), или сохранившие ее только в молодом возрасте, или полностью ее потерявшие, и т. д. Однако Вайден-райх дает этим фактам иное истолкование — он говорит о приобретении новой реакции и о ее результате, как о новой модификации, причем способность к целесообразным реакциям является основным свойством организмов. При этом предполагается далее утеря старой реакции и наследственное «фиксирование» модифи-кационного изменения. Мы считаем, что все эти «новые» реакции имеют уже свое историческое прошлое, т. е. выработались путем отбора как приспособление к переменным условиям среды и имеют уже вполне выраженную наследственную базу. Во всех этих случаях руководящее значение адаптивной модификации не является, правда, экспериментально доказанным, однако логика заставляет нас считать это весьма вероятным. С другой стороны, для некоторых, и в особенности растительных объектов имеются и экспериментальные доказательства значимости модификацион-ыых изменений в процессе образования локальных форм.
Нередко организм, измененный соответственно локальным условиям среды, при переносе в исходные условия возвращается к прежнему фенотипу. Это доказывает, что данное изменение имеет характер модификации (Турессон называет такие локальные формы «экофенами»). Однако обычно возврат не бывает полным, и это доказывает, что модификационное изменение
Глава IV . Прогрессивная эволюция. Адаптациогенез 149
частично или в известных признаках стало наследственным (т. е. было предметом естественного отбора). Таких фактов неполного возврата к исходной форме известно очень много.
Обыкновенная пастушья сумка Capsella bursa pastoris имеет в горах Эрдшиасдаг в Малой Азии типичный альпийский характер — низкий стебель, глубокие корни, ксерофильный облик с опушением. Это растение, по Цедербауеру [Zederbauer, 1908; цит. по: Weidenreich, 1921], могло быть занесено сюда только человеком и, следовательно, лишь недавно стало альпийским растением. При высеве семян в низменности оно частично вернулось к исходной форме — корни и листья потеряли альпийский характер. Однако оно сохранило (в течение более четырех поколений) низкий стебель. С другой стороны, сеянцы обыкновенной равнинной формы пастушьей сумки дают в горах форму, по фенотипу идентичную с горной. Следовательно, весь этот альпийский облик горного растения есть действительно результат модификации обычной пастушьей сумки.
Чистые линии льна, высеянные в разных широтах, дают фе-нотипические изменения, сходные с географическими формами данных мест. Северный лен-долгунец отличается большей высотой и меньшим числом коробочек, чем южный лен-кудряш. На юге лен-долгунец становится ниже, и число стеблей и коробочек увеличивается. На севере лен-кудряш приближается к долгунцу. Следовательно, и здесь формирование географических рас льна шло по пути модификационных изменений. Эти расы дают еще частичный возврат к некоторой исходной форме при изменении климатических условий.
Иногда, однако, частичный возврат к исходным формам при восстановлении прежних условий возможен и после весьма длительной эволюции организма в новой среде. Известно, что многие амфибиотические растения дают, в зависимости от воздушной или водной среды обитания, два рода листьев — наземные и водные (водяной лютик, водяная гречиха, стрелолист и мн. др.). От таких амфибиотических форм произошли все высшие водные растения, у которых, конечно, развиваются только водные листья. На суше такие растения просто гибнут (наземная реакция полностью утеряна), однако, по исследованиям Н. Вакуленко [1940], при прорастании зимних почек типичного водяного растения Myriophyllum verticillatum в воздушной среде (во влажной камере) на верхних частях стебля развивались еще целытокрай-ние листья.
Можно взять также примеры и из области зоологии. И здесь мы знаем немало фактов, показывающих влияние климата в смысле модификационных изменений, в особенности в отношении Животных, обладающих широким географическим распространением. Здесь нередко представители определенного вида образуют северные и южные модификации, отличающиеся очень часто по
150 Организм как целое в индивидуальном и историческом развитии .
размерам, по густоте шерстного покрова (млекопитающие) или по пигментации (бабочки, птицы). Нередко такие модификации имеют явно адаптивное значение (густота шерстного покрова, длина хвоста и ушей, играющих у грызунов роль терморегулятора), и в этом случае они, конечно, выработались как наследственно обусловленные реакции в течение предыдущей истории этих животных. Совершенно подобные же признаки адаптивного, по-видимому, значения образуют наследственную характеристику различных северных и южных видов птиц и млекопитающих (яркая пигментация тропических птиц, короткие уши и хвосты грызунов северных стран). Существование этого «параллелизма» модификационных и видовых изменений ламаркисты считают доказательством наследования «приобретенных» признаков. В самом же деле при оседании нового вида в определенной местности он утрачивает свои излишние адаптивные реакции (например, северный грызун теряет способность развивать более длинный хвост и уши при перенесении в среду с более высокой температурой) и потому кажется приобретшим более прочную наследственную структуру.
Дальнейшим примером может служить переход некоторых хвостатых амфибий к постоянной жизни в воде. Иногда это наблюдается в виде индивидуальной особенности обычно наземных форм (случайная неотения), иногда становится почти постоянным признаком (аксолотль). Способность превращаться во взрослую форму может еще сохраняться (амблистома), но в дальнейшим теряется (постоянножаберные: менобранх, протей и др.). Живородящая альпийская саламандра (Salamandra atra) рождает уже превращенных молодых саламандр; однако у зародышей жабры развиваются и могут еще функционировать как таковые при преждевременном освобождении личинок из тела матери. При этом они приобретают типичную структуру водных жабер. Данная реакция еще не утрачена.
Чаще всего и у животных наблюдается лишь частичный возврат к прежнему фенотипу при восстановлении прежних условий. В этих случаях прежние реакции еще полностью не утеряны. Таковы известные эксперименты воспитания молодых камбал в освещенном снизу аквариуме, в результате чего частично восстанавливалась пигментация нижней стороны их тела [Cunningham, 1891]. Пустынный ткач Австралии (Munia flaviprumna) с однотонной желтой окраской при трехлетнем содержании во влажном климате в Англии получил темпую пигментацию, приближающуюся к другому австралийскому ткачу — Munia castaneithorax, который не живет в пустынях. Очевидно, здесь имел место частичный возврат к исходной окраске близкого вида [Seth-Smith; цит. по: Hosse, 1924]. Таково же значение некоторых опытов Каммерера, получившего восстановление утраченных инстинктов у жабы-повитухи при насильственном удерживании ее в воде в
Глава IV . Прогрессивная эволюция. Адаптациогенез 151
период размножения, как частичный возврат к исходной форме размножения путем откладывания икры в воду. Тот же Каммерер получил и экспериментальный возврат к исходной форме размножения у альпийской саламандры. О том же говорит и возможность частичного восстановления органа зрения и пигментации кожи у протея при воспитании его личинок на свету.
Исследования Юзбашьяна [1940] показали, что при содержании совершенно слепых пещерных креветок (Troglocaris Sclimid-ti Kutaissiana) на свету в аквариуме у многих из них через 72 — 17г года в большей или меньшей мере восстанавливается красный пигмент на дистальных концах глазных стебельков. И здесь, следовательно, частично сказывается роль модификацион-ных изменений в процессе редукции глаз.
Можно еще взять в качестве примера из области морфологии животных явления модифицирующего влияния функции. Предположим, мы имеем перед собой животное с сильно развитой жевательной мускулатурой и с соответственно сильно развитыми гребнями на черепе, служащими для прикрепления этой мускулатуры. Такое строение челюстей, мускулатуры и гребней на черепе имеет свое историческое обоснование и является, очевидно, наследственным. Однако эти соотношения легко модифицируются под влиянием функции. Делались эксперименты с перерезкой нервов, снабжающих жевательную мускулатуру, на одной стороне головы у молодого животного мышцы были здесь парализованы и гребни на черепе недоразвивались.
Подобные гребни имеются на черепе и у человекообразных обезьян. И здесь их развитие определяется, очевидно, развитием мышц; по крайней мере у молодого животного их нет, и у самок при более слабом развитии челюстей и гребни на черепе гораздо менее заметны (рис. 19, 20, 21). Мы, конечно, не можем утверждать, что у предков человека на черепе имелись такие гребни, но все же это возможно. При переходе к иной жизни, когда возрастало значение рук, челюсти уменьшились, жевательная мускулатура становилась слабее и гребни на черепе исчезали. Все это были модификационные изменения. Однако не может быть сомнения, что и сама способность образовать гребень теряется, и как бы человек ни упражнял свою жевательную мускулатуру, у него на черепе гребни не вырастут. Таким образом, может показаться, что модификационное изменение, развившееся вследствие «неупотребления», превратилось в наследственное. На самом деле эта модификация имела и раньше (у человекообразной обезьяны) такую же наследственную базу, как и теперь у человека. Выпало необходимое для развития гребней функциональное раздражение, и морфообразовательная реакция (развитие гребней) перестала осуществляться. Вместе с тем постепенно, в течение многих поколений все более распадался наследственный механизм, контролировавший развитие гребней на черепе,
152 Организм как целое в индивидуальном и историческом развитии
и способность образовать гребни была, наконец, окончательно потеряна (рис. 22).
Таким образом, кажущееся наследственное фиксирование мо-дификационных изменений объясняется лишь потерей излишних реакционных механизмов при переходе организма в иную среду. Естественный отбор не только непрерывно создает новые реакционные и корреляционные механизмы, обеспечивающие нормальное развитие прогрессирующего организма, по и поддерживает существование всех таких механизмов до тех пор, пока они не утратят своего значения в силу изменения соотношений организма со средой (будь то вследствие изменения среды или вследствие изменения самого организма). Все реакционные механизмы, утратившие свое значение, неизменно разрушаются вследствие прекращения естественного отбора по данным механизмам, приводящего к беспорядочному накоплению мутационных изменений. Разрушение этих наследственных систем связано всегда с
|
|
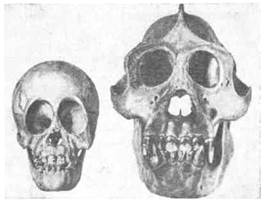
Рис. 19. Черепа детеныша и взрослого самца орангутанга. Развитие гребней черепа [Моллисон, 1933]
Рис. 20. Череп взрослой самки гориллы [Моллисон, 1933]
Рис. 21. Череп взрослого самца гориллы [Моллисон, 1933]
|
|

| Глава IV . Прогрессивная эволюция. Адаптациогенез 153 |

недоразвитием известных частей или с невыявлением структурных и функциональных особенностей. Если эти реакционные системы были связаны с факторами внешней среды, то их выпадение с изменением последней производит ложное впечатление наследственного закрепления модификационного изменения.
Дата добавления: 2019-01-14; просмотров: 275; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!