Малюнок 1. Динаміка гормонів в організмі корів на протязі статевого циклу.
Циклічний характер репродуктивних процесів у самок значною мірою визначається статевим диференціюванням гіпоталамуса по жіночому типу.
Статеве диференціювання гіпоталамуса відбувається під впливом коротко часового утворення та дії статевих стероїдів на ранніх етапах онтогенезу.
В гіпоталамусі самок функціонують циклічний та статичний центри. Їх сумісна діяльність і визначає періодичність в утворенні гонадотропінів базофільними клітинами аденогіпофіза та надходження їх в кров (Гришко Д.С., 2003; Яблонський В.А., 1995; Бакл Д., 1986).
Проте циклічний характер функціонування репродуктивної системи самок обумовлений лише частково жіночим типом діяльності гіпоталамічних центрів. Останній визначає лише потенціальну можливість циклічних процесів. Реалізація цієї можливості відбувається в результаті хвилеподібного характеру росту і визрівання фолікулів і жовтих тіл в яєчниках, які утворюють статеві стероїди – естрогени, прогестерон та андрогени під регулюючим впливом гіпоталамо-гіпофізарної системи.

Фолікуло- та лютеогенез в яєчниках корови представлений на малюнках 2, 3, 4, 5, 6, 7, 8, 9, 10, 11. яєчники зображені в різних масштабах та після сканування.
Малюнок 2. Яєчник корови на другий день після овуляції.

Малюнок 3. Яєчник корови на четвертий день після овуляції.
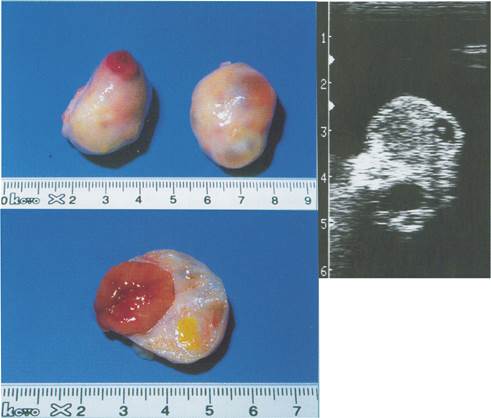
Малюнок 4. Яєчник корови на п’ятий день після овуляції.

Малюнок 5. Яєчник корови на шостий день після овуляції.
|
|
|
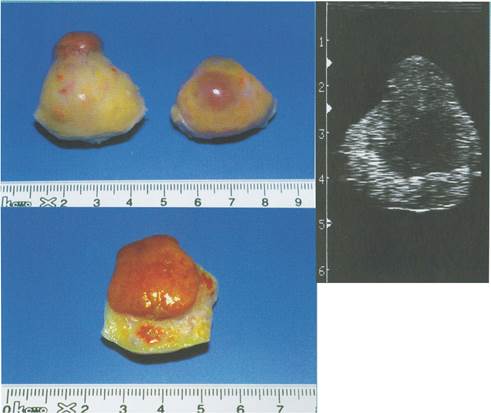
Малюнок 6. Яєчник корови на сьомий день після овуляції.

Малюнок 7. Яєчник корови на дев’ятий день після овуляції.
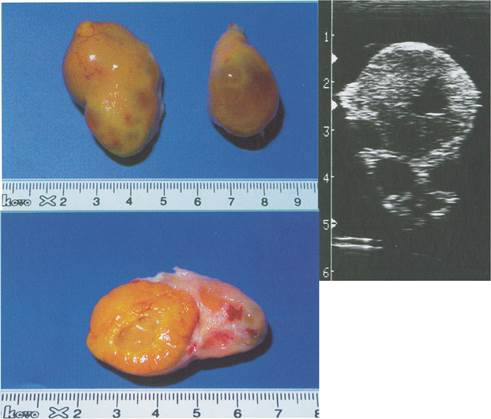
Малюнок 8. Яєчник корови на 15 день після овуляції.
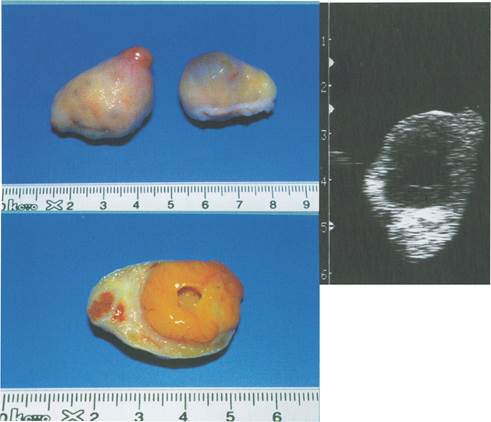
Малюнок 9. Яєчник корови на 16 день після овуляції.

Малюнок 10. Яєчник корови на 17 день після овуляції.

Малюнок 11. Яєчник корови на 20 день після овуляції.
Періодичність продукції цих гормонів (перш за все естрогенів та прогестерона), що залежить від хвилеподібно змінюючогося функціонального стану яєчників, справляє прямий вплив не тільки на статевий апарат, а й зворотній на функціонування центрів гіпоталамуса. Циклічні зміни в рівнях секреції різних оваріальних гормонів і співвідношення цих рівнів за механізмом зовнішнього зворотнього зв’язку (послідовного чергування позитивної та негативної) періодично модулює функціональний стан обох гіпоталамічних центрів і тим самим підтримує їх циклічну діяльність. Малі дози естрогенів і прогестинів модулюють, а великі блокують активність центрів гіпоталамуса і функції гіпофіза.
Таким чином циклічність статевих процесів у самок обумовлена двома рядами взаємопов’язаних механізмів: жіночим типом функціонально-структурної організації гіпоталамічної регуляції, що включає два центри контролю гонадотропних функцій гіпофіза, чутливих до змін рівней оваріальних гормонів; періодичністю діяльності яєчників, гормони яких здатні ритмічно змінювати функціональний стан обох центрів гіпоталамуса і внаслідок цього гонадотропної та інших функцій гіпофіза.
|
|
|
Тільки спільне, точне, збалансоване функціонування цих двох механізмів та цілісність гіпоталамо-гіпофізарної системи можуть забезпечити реалізацію та нормальний перебіг статевих циклів. Відкриття нейросекреції помогло подолати розрив між нервовою та ендокринною системами, який існував в фізіології на протязі багатьох десятків років. Стало зрозумілим, що нейроендокринна система забезпечує єдність нервової та гормональної регуляції в організмі. Межі цієї системи, яка спочатку включає лише гіпоталамус, зараз значно розширились і увібрали в себе епіфіз, кінцеву пластину, гіпокамп, мігдалевидні ядра та інші структури мозку. Сукупність нейросекреторних елементів деколи називають „ендокринним мозком” – поняття не тільки образне, але й достатньо точне та ємке.
Утворення та (або) секреція аденогіпофізарних гормонів знаходиться під подвійним контролем з боку гіпоталамуса. По гіпоталамо-гіпофізарній портальній системі в аденогіпофіз надходять ліберіни та статини і баланс між цими факторами визначає секрецію аденогіпофізарних гормонів в кожний даний момент. Фолікулостимулюючий (ФСГ) і лютеінізуючий (ЛГ) гормони відносяться до гонадотропних, тобто вони безпосередньо приймають участь в регуляції активності гонад (Недвига О., 2003; Павлов В.А., 1984).
|
|
|
Таким чином з врахуванням вищенаведеного, потрібно зробити висновок: гіпофіз не є „головна” залоза („диригент”) внутрішньої секреції, яка контролює всі інші залози, але цілком очевидно, що це дуже важливий регуляторний центр, який об’єднує нервовий та ендокринний компоненти координуючої системи організму.
Гістологічним та гістохімічним дослідженнями показано, що в нормі в аденогіпофізі присутні загальний білок, ДНК, глікоген. Активно функціонуючі аденоцити не бувають вільні від секреторних гранул. Тучні клітини включаються в гомеостаз органу та можуть слугувати побічним показником активності функціонування залози.
Висловлюється думка (Павлов В.А., 1984), що статева циклічність у тварин може бути порушена внаслідок зниженої чутливості гіпоталамуса до естрогенів, недостатньої активності гіпофіза і, що в основі всіх форм неплідності, викликаних порушеннями функції яєчників та матки, лежать ендокринні розладнання гіпоталамо-гіпофізарної системи, щитовидної залози, наднирників і яєчників. Проте питання ці вкрай недостатньо вивчені.
|
|
|
Нами (Кошовий В.П., 1981) в експериментах з’ясований вплив вітаміну А на структуру та функцію аденогіпофіза телиць. Так, при дефіциті вітаміну А в аденогіпофізі виявлені значні морфологічні та функціональні зміни. Ці зміни характеризуються наявністю глибоких дистрофічних процесів „мілкокістозної” дистрофії, десквамації базофілів і ацідофілів, дегрануляції цих клітин. Триразове (інтервал 6 – 7 днів, доза 1 млн і.о. на введення) застосування ретинолу ацетату телицям з дефіцитом вітаміну А підвищує у них (на 67,9%. Р < 0,001) кількість базофілів і ацідофілів та вміст в плазмі цих клітин РНК. У таких телиць відсутні дистрофічні процеси в аденогіпофізі. Все це пояснює підвищення гонадотропної активності аденогіпофіза під впливом вітаміну А (малюнки 12, 13).

Дата добавления: 2018-11-24; просмотров: 689; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!