Головной мозг человека
Головной мозг — важнейший орган, координирующий и регулирующий все жизненные функции организма и контролирующий поведение. Все наши мысли, чувства, ощущения, желания и движения связаны с работой мозга, и если он не функционирует, человек переходит в вегетативное состояние: утрачивается способность к каким-либо действиям, ощущениям или реакциям на внешние воздействия. Мозг человека более сложный и высокоорганизованный, чем мозг животных. Однако существует значительное сходство в устройстве мозга человека и других млекопитающих, как, впрочем, и большинства видов позвоночных. Центральная нервная система состоит из головного и спинного мозга. Она связана с различными частями тела периферическими нервами — двигательными и чувствительными.
Головной мозг — симметричная структура, как и большинство других частей тела. При рождении ребенка вес головного мозга составляет примерно 0,3 кг, тогда как у взрослого — около 1,5 кг. При внешнем осмотре мозга выделяют два больших полушария, скрывающие под собой глубинные образования. Поверхность полушарий покрыта бороздами и извилинами, увеличивающими поверхность коры (наружного слоя мозга). Сзади помещается мозжечок, поверхность которого более тонко изрезана. Ниже больших полушарий расположен ствол мозга, переходящий в спинной мозг. От ствола и спинного мозга отходят нервы, по которым к мозгу стекается информация от внутренних и наружных рецепторов, а в обратном направлении идут сигналы к мышцам и железам. От головного мозга отходят 12 пар черепно-мозговых нервов.
|
|
|
Внутри мозга различают серое вещество, состоящее преимущественно из тел нервных клеток и образующее кору, и белое вещество — нервные волокна, которые формируют проводящие пути (тракты), связывающие между собой различные отделы мозга, а также образуют нервы, выходящие за пределы ЦНС и идущие к различным органам.
Головной и спинной мозг защищены костными футлярами — черепом и позвоночником. Между веществом мозга и костными стенками располагаются три оболочки: наружная — твердая мозговая оболочка, внутренняя — мягкая, а между ними — тонкая паутинная оболочка. Пространство между оболочками заполнено спинномозговой (цереброспинальной) жидкостью, которая по составу сходна с плазмой крови, вырабатывается во внутримозговых полостях (желудочках мозга) и циркулирует в головном и спинном мозгу, снабжая его питательными веществами и другими необходимыми для жизнедеятельности факторами.
Кровоснабжение головного мозга обеспечивают в первую очередь сонные артерии; у основания мозга они разделяются на крупные ветви, идущие к различным его отделам. Хотя вес мозга составляет всего 2,5 % веса тела, к нему постоянно, днем и ночью, поступает 20% циркулирующей в организме крови и соответственно кислорода. Энергетические запасы самого мозга крайне невелики, так что он чрезвычайно зависим от снабжения кислородом. Существуют защитные механизмы, способные поддержать мозговой кровоток в случае кровотечения или травмы. Особенностью мозгового кровообращения является также наличие так называемого гематоэнцефалического барьера. Он состоит из нескольких мембран, ограничивающих проницаемость сосудистых стенок и поступление многих соединений из крови в вещество мозга; таким образом, этот барьер выполняет защитные функции. Через него не проникают, например, многие лекарственные вещества.
|
|
|
Клетки ЦНС называются нейронами; их функция — обработка информации. В мозгу человека от 5 до 20 млрд нейронов. В состав мозга входят также глиальные клетки, их примерно в 10 раз больше, чем нейронов. Глия заполняет пространство между нейронами, образуя несущий каркас нервной ткани, а также выполняет метаболические и другие функции.
Нейрон, как и все другие клетки, окружен полупроницаемой (плазматической) мембраной. От тела клетки отходят два типа отростков — дендриты и аксоны. У большинства нейронов много ветвящихся дендритов, но лишь один аксон. Дендриты обычно очень короткие, тогда как длина аксона колеблется от нескольких сантиметров до нескольких метров. Тело нейрона содержит ядро и другие органеллы, такие же, как и в других клетках тела.
|
|
|
Нейромедиатор, выделившийся из пресинаптической мембраны аксона, связывается с рецепторами на дендритах постсинаптического нейрона. Мозг использует разнообразные нейромедиаторы, каждый из которых связывается со своим особым рецептором.
С рецепторами на дендритах соединены каналы в полупроницаемой постсинаптической мембране, которые контролируют движение ионов через мембрану. В покое нейрон обладает электрическим потенциалом в 70 мВ (потенциал покоя), при этом внутренняя сторона мембраны заряжена отрицательно по отношению к наружной. Хотя существуют различные медиаторы, все они оказывают на постсинаптический нейрон либо возбуждающее, либо тормозное действие. Возбуждающее влияние реализуется через усиление потока определенных ионов, главным образом натрия и калия, через мембрану. В результате отрицательный заряд внутренней поверхности уменьшается — происходит деполяризация. Тормозное влияние осуществляется в основном через изменение потока калия и хлоридов, в результате отрицательный заряд внутренней поверхности становится больше, чем в состоянии покоя, и происходит гиперполяризация.
|
|
|
Миелин и глиальные клетки. Многие аксоны покрыты миелиновой оболочкой, которая образована многократно закрученной мембраной глиальных клеток. Миелин состоит преимущественно из липидов, что и придает характерный вид белому веществу головного и спинного мозга. Благодаря миелиновой оболочке скорость проведения потенциала действия по аксону увеличивается, так как ионы могут перемещаться через мембрану аксона лишь в местах, не покрытых миелином, — так называемых перехватах Ранвье. Между перехватами импульсы проводятся по миелиновой оболочке как по электрическому кабелю. Поскольку открытие канала и прохождение по нему ионов занимает какое-то время, устранение постоянного открывания каналов и ограничение их сферы действия небольшими зонами мембраны, не покрытыми миелином, ускоряет проведение импульсов по аксону примерно в 10 раз. Только часть глиальных клеток участвует в формировании миелиновой оболочки нервов (шванновские клетки) или нервных трактов (олигодендроциты). Гораздо более многочисленные глиальные клетки (астроциты, микроглиоциты) выполняют иные функции: образуют несущий каркас нервной ткани, обеспечивают ее метаболические потребности и восстановление после травм и инфекций.
Рассмотрим простой пример. Что происходит, когда мы берем в руку карандаш, лежащий на столе? Свет, отраженный от карандаша, фокусируется в глазу хрусталиком и направляется на сетчатку, где возникает изображение карандаша; оно воспринимается соответствующими клетками, от которых сигнал идет в основные чувствительные передающие ядра головного мозга, расположенные в таламусе (зрительном бугре), преимущественно в той его части, которую называют латеральным коленчатым телом. Там активируются многочисленные нейроны, которые реагируют на распределение света и темноты. Аксоны нейронов латерального коленчатого тела идут к первичной зрительной коре, расположенной в затылочной доле больших полушарий. Импульсы, пришедшие из таламуса в эту часть коры, преобразуются в ней в сложную последовательность разрядов корковых нейронов, одни из которых реагируют на границу между карандашом и столом, другие — на углы в изображении карандаша и т. д. Из первичной зрительной коры информация по аксонам поступает в ассоциативную зрительную кору, где происходит распознавание образов, в данном случае карандаша. Распознавание в этой части коры основано на предварительно накопленных знаниях о внешних очертаниях предметов.
Планирование движения (т. е. взятия карандаша) происходит, вероятно, в коре лобных долей больших полушарий. В этой же области коры расположены двигательные нейроны, которые отдают команды мышцам руки и пальцев. Приближение руки к карандашу контролируется зрительной системой и интерорецепторами, воспринимающими положение мышц и суставов, информация от которых поступает в ЦНС. Когда мы берем карандаш в руку, рецепторы в кончиках пальцев, воспринимающие давление, сообщают, хорошо ли пальцы обхватили карандаш и каким должно быть усилие, чтобы его удержать. Если мы захотим написать карандашом свое имя, потребуется активация другой хранящейся в мозге информации, обеспечивающей это более сложное движение, а зрительный контроль будет способствовать повышению его точности.
На приведенном примере видно, что выполнение довольно простого действия заставляет работать обширные области мозга, простирающиеся от коры до подкорковых отделов. При более сложных формах поведения, связанных с речью или мышлением, активируются другие нейронные цепи, охватывающие еще более обширные области мозга.
Другие системы. Лимбическая система — широкая сеть связанных между собой областей мозга, которые регулируют эмоциональные состояния, а также обеспечивают научение и память. К ядрам, образующим лимбическую систему, относятся миндалевидные тела и гиппокамп (входящие в состав височной доли), а также гипоталамус и ядра так называемой прозрачной перегородки (расположенные в подкорковых отделах мозга).
Ретикулярная формация — сеть нейронов, протянувшаяся через весь ствол к таламусу и далее связанная с обширными областями коры. Она участвует в регуляции сна и бодрствования, поддерживает активное состояние коры головного мозга и способствует фокусированию внимания на определенных объектах.
Биометрия
С помощью электродов, размещенных на поверхности головы или введенных в вещество мозга, можно зафиксировать электрическую активность мозга, обусловленную разрядами его клеток. Запись электрической активности мозга с помощью электродов на поверхности головы называется электроэнцефалограммой (ЭЭГ). Она не позволяет записать разряд отдельного нейрона. Только в результате синхронизированной активности тысяч или миллионов нейронов появляются заметные колебания (волны) на записываемой кривой. При постоянной регистрации на ЭЭГ выявляются циклические изменения, отражающие общий уровень активности индивида. В состоянии активного бодрствования ЭЭГ фиксирует низкоамплитудные неритмичные бета-волны. В состоянии расслабленного бодрствования с закрытыми глазами преобладают альфа-волны частотой 7–12 циклов в секунду. О наступлении сна свидетельствует появление высокоамплитудных медленных волн (дельта-волн). В периоды сна со сновидениями на ЭЭГ вновь появляются бета-волны, и на основании ЭЭГ может создаться ложное впечатление, что человек бодрствует (отсюда термин “парадоксальный сон”). Сновидения часто сопровождаются быстрыми движениями глаз (при закрытых веках). Поэтому сон со сновидениями называют также сном с быстрыми движениями глаз. ЭЭГ позволяет диагностировать некоторые заболевания мозга, в частности эпилепсию.
Если регистрировать электрическую активность мозга во время действия определенного стимула (зрительного, слухового или тактильного), то можно выявить так называемые вызванные потенциалы — синхронные разряды определенной группы нейронов, возникающие в ответ на специфический внешний стимул. Исследование вызванных потенциалов позволило уточнить локализацию мозговых функций, в частности связать функцию речи с определенными зонами височной и лобной долей. Это исследование помогает также оценить состояние сенсорных систем у больных с нарушением чувствительности.
К числу важных нейромедиаторов мозга относятся ацетилхолин, норадреналин, серотонин, дофамин, глутамат, гамма-аминомасляная кислота (ГАМК), эндорфины и энкефалины. Помимо этих хорошо известных веществ в мозге, вероятно, функционирует большое количество других, пока не изученных. Некоторые нейромедиаторы действуют только в определенных областях мозга. Так, эндорфины и энкефалины обнаружены лишь в путях, проводящих болевые импульсы. Другие медиаторы, такие, как глутамат или ГАМК, более широко распространены.
Действие нейромедиаторов. Как уже отмечалось, нейромедиаторы, воздействуя на постсинаптическую мембрану, изменяют ее проводимость для ионов. Часто это происходит через активацию в постсинаптическом нейроне системы второго “посредника”, например циклического аденозинмонофосфата (цАМФ). Действие нейромедиаторов может видоизменяться под влиянием другого класса нейрохимических веществ — пептидных нейромодуляторов. Высвобождаемые пресинаптической мембраной одновременно с медиатором, они обладают способностью усиливать или иным образом изменять эффект медиаторов на постсинаптическую мембрану.
Большое значение имеет недавно открытая эндорфин-энкефалиновая система. Энкефалины и эндорфины — небольшие пептиды, которые тормозят проведение болевых импульсов, связываясь с рецепторами в ЦНС, в том числе в высших зонах коры. Это семейство нейромедиаторов подавляет субъективное восприятие боли.
Электрохимия и электрические процессы, протекающие в живом организме, тесно связаны с информационными вопросами понимания так называемого “энергетического состояния”. Изучение энергетических процессов в живом организме строится на известных законах преобразования энергии из одного вида в другой. Можно сказать, что источники энергии определяют принцип существования живых организмов в Природе. В современном представлении все живые организмы можно представить в виде двух групп в зависимости от того, в какой химической форме они получают углерод из окружающей среды.
Автотрофы (от греческого авто — само, трофа — пища) представляют собой группу самостоятельно питающихся организмов, которые могут использовать в качестве единственного источника углерода — оксид углерода СО2. К автотрофам относят растения фотосинтезирующие и хемосинтезирующие бактерии.
Гетеротрофы (от греческого гетерос — другой, трофа — пища) представляют собой группу организмов, которые получают углерод в виде готовых достаточно сложных органических соединений, например углеводов. К этой категории относятся животные и большое количество микроорганизмов.
Здесь важно отметить, что для энергетических процессов важным компонентом является использование определенных биохимических соединений в качестве либо донора, либо акцептора электронов. Из нашего практического опыта мы знаем, что многие организмы могут существовать как в аэробных, так и анаэробных условиях. В аэробных условиях они используют в качестве акцепторов кислород, который поступает в организм за счет дыхания. В анаэробных же условиях акцепторами электронов являются органические вещества, на основе которых работает механизм “брожения”. Этот механизм создания запасов энергии — преобразования энергии — не очень эффективен, но его реализация в Природе имеет очень длинную историю. И это действительно так, потому, что весь кислород, находящийся в атмосфере Земли, образовался в процессе фотосинтеза, другими словами, за счет деятельности анаэробной энергетики.
По современной терминологии высокоэнергетическими соединениями называются АТФ и вещества, способные образовывать АТФ в ферментативных реакциях переноса групп без участия окислительных процессов. Такие соединения содержат в своей молекуле связи, при гидролизе которых высвобождается большое количество свободной энергии. Реакция протекает при различных условиях, оказывающих влияние на величину изменения свободной энергии. Поэтому в биохимии используют термин — изменение стандартной свободной энергии — D G0 . Этим термином обозначают такое изменение свободной энергии при стандартных условиях: давлении в 1 атм.; исходной концентрации субстратов — 1 М; температуре — 25 ° C; pH = 7,0. Значение D G0 применяют для количественной характеристики как метаболических процессов, так и некоторых химических реакций. Такая интерпретация D G0 помогает соединить биохимические и термодинамические представления о механизме преобразования и расходования энергии в живом организме. Как и в термодинамике, здесь знаком “минус” обозначается отдача энергии, а знаком “плюс” ее приобретение. Если текущее значение D G0 для некоторой реакции имеет отрицательное значение, то принято считать, что такая реакция может происходить самопроизвольно с выделением свободной энергии. Такие реакции носят название экзергонических. Напротив, если текущее значение D G0 для некоторой реакции имеет положительное значение, то такая реакция происходит с поглощением энергии. Такие реакции носят название эндергонических. В качестве посредника между процессами, связанными с генерированием и использованием энергии, функционирует система высоко энергетических химических соединений. Величина стандартной свободной энергии гидролиза высокоэнергетических связей превышает 21 кДж/моль.
Высокоэнергетические соединения имеют, как правило, в своем составе высокоэнергетическую фосфатную группу и могут передавать ее другим веществам. Различают пять основных типов высокоэнергетических соединений: рибонклеозид 5’ — дифосфаты и трифосфаты; карбоксилфосфаты; ацилтиоловые эфиры; фосфорамидные соединения; енолфосфаты. Принимая во внимание, что вся энергетика живого организма строится по подобию клеточных энергетических процессов, выделяют аденилатную систему: АТФ и продукты ее гидролиза. Такая система может быть в самой простой образной форме представлена в виде “аккумулятора”, который постоянно заряжается энергией от других генераторов, и одновременно с этим снабжает энергией множество клеточных систем и тканей живого организма. И тут, можно сказать, мы подходим к главному пониманию энергетического процесса. Весь процесс “зарядки аккумулятора” представляется синтезом АТФ:
АДФ + Фи ® Е1 ® АТФ + Н2О, (1)
а “разрядка аккумулятора” сопровождается гидролизом АТФ:
АТФ + Н2О ® Е2 ® АДФ + Фи, (2)
где Фи можно для простоты рассмотрения представить как продукт гидролиза АТФ, а Е1 и Е2 — отождествлять с некоторым набором ферментов, при наличии которых данные реакции могут быть осуществлены.
Рассматривая полученные результаты, отмечаем их компактность представления об основной концепции жизни — наличии диссипативной системы. В такой открытой системе, далекой от равновесия, возникает динамическая упорядоченность. Полагая, что такая система отождествляется с живым организмом, мы ожидаем увидеть когерентное поведение, которое, как мы уже знаем, отвечает условию гомеостазиса. Наряду с этим не будем отклоняться от понимания прямого и обратного процессов, свидетельствующих о необходимости учитывать их неравнозначность, некоторую квазипериодичность и одновременно с этим очень сильную взаимосвязанность. В основе теории биологических систем так, как это мы представляем сегодня, на основах непостоянства, неточного соответствия прямого и обратного процессов рождается живая ткань. И здесь становится очевидным, что критерием возможности возникновения динамической упорядоченности в диссипативной системе может служить уровень отклонения от равновесного состояния.
Как мы отмечали, характеристика сложной биохимической реакции, состоящей из многих этапов, может быть выражена в типовых энергетических величинах, например в джоулях или калориях. Мы можем несколько расширить наши представления о живом организме и по аналогии с предыдущим сказать, что живая система в определенном смысле машина по преобразованию энергии. В такой системе в аналогичных единицах выражается энтропийная характеристика. В свою очередь, энтропия связана с информацией. Не включая всего механизма определений этих понятий, которые в той или иной степени должны быть знакомы читателю, рассмотрим тот аспект проблемы взаимосвязи биологических процессов с термодинамикой, который может быть раскрыт на представлениях об информационном потоке. На этой общей ступени рассмотрим простой пример, который в некоторой степени прояснит направленность наших рассуждений.
Будем исходить из того, что существуют P 0 равновероятных событий, которые могут произойти в некоторой системе. Если на известном интервале времени T происходит Р 1 событий, то информация о поведении системы, исчисляемая в битах, вычисляется так:
I = k ln P 0 / P 1 = k lg P 0 – k lg P 1,
где k — постоянная, зависящая от выбора системы единиц. Целесообразность представления информации через логарифмическую функцию продиктована желанием подчеркнуть свойство аддитивности информационного потока. Применительно ко многим задачам биометрии допустимо полагать значение k = 2,078, а само определение количества информации вести по известной формуле Шеннона:
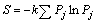 , (3)
, (3)
которая может быть охарактеризована как информационная энтропия. С точки зрения эволюционного процесса в системе, нас интересует характер изменения информации, который мы смогли бы наблюдать при смене одного состояния системы на другое. Такой информационный индекс можно определить так:
 . (4)
. (4)
Для задач, отдаленно связанных с биометрией, рекомендуется пользоваться постоянной Больцмана (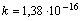 Дж/К).
Дж/К).
Для пояснения полученного соотношения рассмотрим такую задачу. Положим, имеется некоторая система, которая самостоятельно способна перейти из состояния Р = (Р 1, Р 2, …, Рn) в состояние Q = (Q 1, Q 2, …, Qn). Возникает естественный вопрос о том, как будет изменяться информация о системе. Очевидно, что при пошаговом переходе от Р к Q справедливо равенство
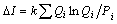 .
.
Информационный индекс, при таком процессе всегда положителен. При Q = P сразу получается D I = 0. Из этого можно сделать вывод о том, что знание о “переходе” системы из Р в состояние Q уменьшает неопределенность и повышает информационную составляющую. Здесь мы приходим к выводу, что информационная энтропия тесно связана с реальной физической энтропией системы. С точки зрения практики это означает, что за полученную в ходе исследования объекта информацию надо платить увеличением энтропии окружающей среды. В таком случае, а это уже сегодня стало одним из биологических постулатов, мы можем утверждать, что сумма информации и энтропии всегда величина постоянная:
I + S = const. (5)
Соотношение этих параметров определяет основные динамические характеристики биологической системы. Знание о том, что это соотношение постоянно меняется, позволяет рассматривать много различных математических моделей описания поведения биологической системы. Одновременно с этим при наличии специальных аппаратных средств измерений появляется возможность зарегистрировать текущие состояния системы, представить их в форме информационных потоков. Регистрация информационных данных о поведении биологической системы, проводимая с помощью технических средств, осуществляется с помощью специальных датчиков.
Из обычных практических наблюдений за некоторыми событиями или явлениями, происходящими в Природе, очень часто получается хорошая концепция понятий и объяснений. Такое положение дел существует в тех областях исследований, где теоретические основы очень тесно переплелись и составили единое поле взаимосвязанных представлений. На основании такого подхода можно показать, что изменение во времени некоторого существенного параметра биологической системы, представленного в форме информационного сообщения регистрирующего датчика, может быть адекватным образом сопоставлено с энергетическим состоянием системы:
D W =- k lg2/D r, (6)
где D r — обобщенный показатель кратности вырождения исследуемой системы. Размерность параметра D W получается в энергетических единицах и характеризует текущее состояние биологической системы.
Таким образом, используя введенные представления о механизмах образования сигнала, об энергетических процессах, протекающих в живом организме и направленных на реализацию закона гомеостазиса, мы пришли к пониманию взаимосвязанности этих многообразных явлений. Так, понятие симметрии и антисимметрии оказалось гораздо более глубоким и нашло свое зеркальное отражение в других не менее важных процессах таких, как “синтез” и “деструкция”, которые представляют собой обобщенный процесс. Понимание механизмов порождения такого процесса может составить основу — ступень, с которой следует приступать к поиску ключа биометрической процедуры, который обязательно должен связывать два нераздельных понятия энтропии и информации.
Начинать, конечно, нужно с энтропии. Это направление хорошо проработано в науке. Для биологии и медицины оно пока остается на уровне простого описания, фиксации уже свершившихся событий. В этих областях знаний еще предстоит научиться строить такие прогнозы, которые хорошо бы согласовывались с наблюдаемыми явлениями. Понятие “энтропийной характеристики” биологического сигнала можно получить, а это сегодня одна из актуальных тем многих научных работ, но вот вопрос о получении универсальной характеристики для разных типов сигналов несколько отодвигает торжества принятия новых постулатов. В биологии, там, где математик или физик заходит на поле проявления особенно ярких свойств Природы, все получается не так гладко, как в обычной физике или химии. Здесь нет возможностей повторить эксперимент в близких, очень близких условиях, и всегда приходится вносить определенные условные допущения. Результат — отсутствие четких законов, постулатов, которые исполняются в широком диапазоне условий эксперимента.
Рассматривая значение энтропии как показатель текущего состояния биологической системы, можно легко подойти к формулировке задачи о биометрии, в терминах которой требуется определить текущее состояние живого организма по данным, выполненным обычными аппаратными методами. Другими словами, мы стремимся провести некоторые натурные измерения на биологической системе с целью узнать ее текущее энергетическое состояние. Полагая, что измерительная процедура дает нам информационную характеристику состояния биологической системы, мы рассчитываем получить энергетическую оценку на основе массива биометрических данных.
Следуя традиционным путем биологической медицинской кибернетики, представим биологическую систему в виде закрытого ящика. Создавая некоторые тестовые воздействия на входе системы, можно убедиться в том, что одна категория возмущений будет приводить к понижению энергетического ресурса, а другая категория воздействий будет увеличивать такой ресурс. Промежуточный случай, когда не будет наблюдаться никаких изменений в системе, теоретически также возможен. Это случай, который может рассматриваться с точки зрения “чувствительности”, развитости внутренней структуры системы, особенностей и продолжительности внутренних информационных циклов.
Дата добавления: 2015-12-21; просмотров: 23; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!