Автотрофное питание
Фотосинтез является одним из двух путей автотроф- ного питания организмов. Характерен для зелёных растений, цианобактерий, зелёных и пурпурных серобактерий, имеющих пигменты: (хлорофилл, фу- коксантин, фикоэритрин, бактериохлорофиллы и др.).
Фотосинтез — процесс первичного синтеза органических веществ из неорганических (углекислого газа и воды) под действием энергии света в присутствии пигментов.
Общее уравнение фотосинтеза: 6Н20 + 6С02 -®->C6Н12O6 + 6O2, где Э означает энергию кванта (фотона) света.
Фотосинтез слагается из двух серий реакций: вызванных светом (световые реакции, или световая фаза) и фиксации углекислого газа (темновые реакции, или темновая фаза).
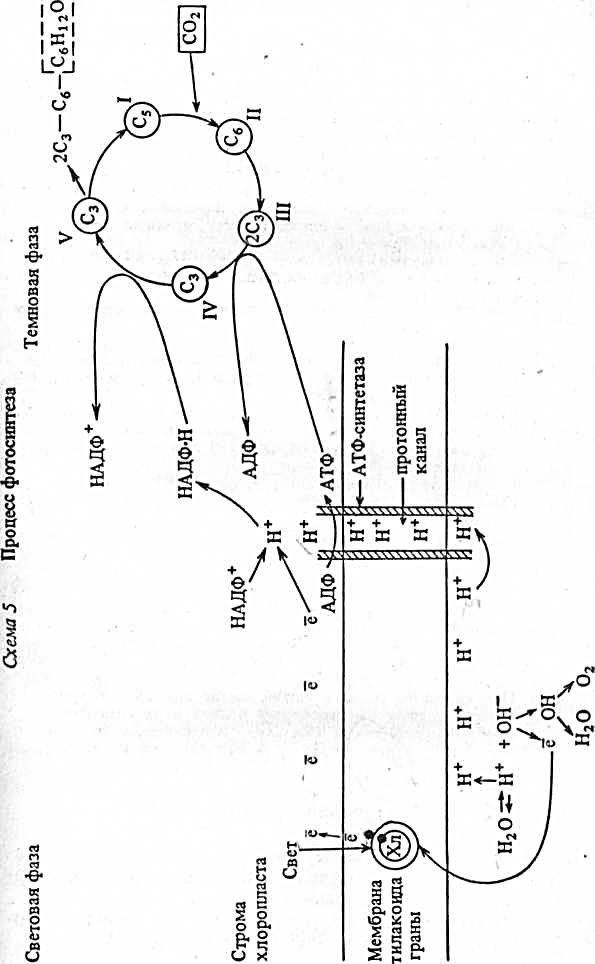
ФАЗЫ ФОТОСИНТЕЗА
1. Световая фаза. Протекает только на свету в гранах тилакоидов хлоропласта.
Она заключается в преобразовании энергии света пигментами, образующими фотосистемы, в энергию химических связей АТФ и одновременно в образовании вещества — переносчика протонов (НАДФ • 2Н), обладающего «восстановительной силой».
Пигменты в гранах образуют фото синтетические единицы — фотосистему I (ФС-1) и фотосистему II (ФС-II). Каждая фотосистема состоит из основного пигмента хлорофилла или (P680 или Р7оо) — реакционного центра и вспомогательных пигментов (200—350 молекул хлорофилла, каротиноидов).
· Квант света, попадая на молекулы хлорофилла, которые находятся в мембрнах тилакоидов гран, приводит их в возбужденное состояние. В результате этого электроны е сходят со своих орбит и переносятся с помощью переносчиков за пределы мембраны тилакоидаде и накапливаются, создавая отрицательно заряженное электрическое поле.
|
|
|
· Место вышедших электронов в молекулах хлорофилла занимают электроны воды ё, так как вода под действием света подвергается фоторазложению (фотолизу): Н20→0Н- + Н+; ОН-→ё→ОН
· Гидроксилы ОН-, став радикалами ОН, объединяются: 40Н→2Н20 + 02, образуя воду и свободный кислород, который выделяется в атмосферу.
· Протоны водорода Н+ не проникают через мембрану тилакоида и накапливаются внутри, образуя положительно заряженное электрическое поле, что приводит к увеличению разности потенциалов по обе стороны мембраны.
· При достижении критической разности потенциалов протоны Н+ устремляются по протонному каналу в ферменте АТФ - синтетаза, встроенному в мембрану тилакоида, наружу. На выходе из протонного канала создается высокий уровень энергии, которая идет на синтез АТФ (АДФ + Ф→АТФ). Образовавшиеся молекулы АТФ переходят в строму, где участвуют в реакциях фиксации углерода.
· Протоны Н+, вышедшие на поверхность мембраны тилакоида, соединяются с электронами ё, образуя атомарный водород Н, который идет на восстановление переносчика НАДФ +;
|
|
|
2ё+Н+ + НАДФ+→НАДФ-Н (переносчик с присоединенным водородом)
Таким образом, активированный световой энергией электрон хлорофилла используется для присоединения водорода к переносчику. НАДФ-Н переходит в строму хлоропласта, где участвует в реакциях фиксации углерода.
2. Темновая фаза. Протекает как на свету, так и в его отсутствие в строме хлоропласта (от наличия света не зависит).
Она заключается в фиксации углекислого газа и синтезе углеводов за счёт энергии АТФ и «восстановительной силы» НАДФ-2Н.
Реакции темновой фазы фотосинтеза были изучены американским ученым М.Кальвиным и названы его именем (цикл Кальвина).
· Реакции фиксации углерода (темновые реакции). Осуществляются в строме хлоропласта, куда поступают АТФ, НАДФ-Н от тилакоидов гран и С02 из воздуха. Кроме того, там постоянно находятся пятиуглеродные соединения — пентозы Сб, которые образуются в цикле Кальвина (цикле фиксации С02). Этот цикл можно проследить на углероде как главном элементе углеводов.
· К пентозе С5 присоединяется С02, в результате чего появляется нестойкое шестиуглеродное соединение С6, которое расщепляется на две трехуглеродные группы 2Сз — триозы.
|
|
|
· Каждая из триоз 2С3 принимает по одной фосфатной группе от 2 АТФ, что обогащает молекулы энергией.
· Каждая из триоз 2С3 присоединяет по одному атому водорода от 2 НАДФ • Н. После чего одни триозы объединяются, образуя углеводы 2Сз→Сб→СбН120б (глюкоза)
· Другие триозы объединяются, образуя пентозы 5Сз→ЗС5, и вновь включаются в цикл фиксации С02.
· Суммарная реакция фотосинтеза: 6С02 + 6Н20→→ С6Н1206 + 602
· 
|
Значение фотосинтеза
· образование в биосфере свободной энергии в виде первичного органического вещества, (ежегодно 150 млрд т), обеспечивающего существование самих растений и гетеротрофных организмов (по мнению К.А. Тимирязева, растения, усваивая солнечную энергию, играют космическую роль);
· выделение в атмосферу кислорода (ежегодно до 200 млрд. т), обеспечивающего клеточное дыхание самих растений и других организмов. Кислород необходим также для образования озона (302 -> 203), образующего озоновый экран, служащий защитой от коротковолнового УФ-излучения;
· получение продовольствия для человека и кормов сельскохозяйственным животным; запасание энергии в виде различных видов топлива (древесина, уголь, торф).
|
|
|
Продуктивность фотосинтеза — скорость образования растениями органических веществ за единицу времени на единицу площади листьев.
| Факторы, влияющие на продуктивность фотосинтеза | ||
| ОСВЕЩЁННОСТЬ | КОНЦЕНТРАЦИЯ С02 | ТЕМПЕРАТУРА |
| При низкой освещённости скорость фотосинтеза прямо пропорциональна интенсивности света. При очень большой освещённости иногда начинается разрушение хлорофилла, и это замедляет фотосинтез. | Повышение концентрации С02 до 0,1-0,3% в условиях тепличного выращивания растений ускоряет скорость фотосинтеза в два раза. | Оптимальная температура для растений умеренного климата около +25 °С. При дальнейшем повышении температуры скорость уменьшается, так как растение тратит много воды на транспирацию (испарение), предотвращающую перегрев. |
Хемосинтез — процесс синтеза некоторыми микроорганизмами органических веществ из неорганических за счёт энергии, получаемой от окисления неорганических соединений: водорода, сероводорода, аммиака и др. Один из двух путей автотрофного питания организмов.
ОСНОВНЫЕ ГРУППЫ ХЕМОСИНТЕЗИРУЮЩИХ БАКТЕРИЙ
| Азотофиксирующие бактерии | Обладают способностью усваивать молекулярный азот воздуха и переводить его в доступные для растений формы. Играют важную роль в круговороте азота в природе. Могут быть аэробами и анаэробами, свободно живущими в почве и живущими в симбиозе с растениями |
| Водородные бактерии | Широко распространены в почве, окисляют водород, постоянно образующийся при анаэробном (бескислородном) разложении различных органических остатков: 2Н2 + 02 -> 2Н20 + энергия.Человек использует водородные бактерии для получения пищевого и кормового белка, полноценного по аминокислотному составу, а также для регенерации атмосферы в замкнутых пространствах |
| Нитрифицирующие бактерии | Живут в почве и осуществляют окисление аммиака, образующегося при гниении органических остатков, до азотной кислоты. Процесс окисления идёт в два этапа (образование нитритов NO2-, затем нитратов NO3-). Участвуют в круговороте азота в природе и таким образом поддерживают плодородие почвы. |
| Серобактерии | Живут в водоёмах, вода которых содержит сероводород. Сероводород окисляют до свободной кристаллической серы. При недостатке сероводорода производят дальнейшее окисление свободной серы до серной кислоты: 2H2S + 02 -> 2HZ0 + 2S + энергия; 2S + 302 + 2Н20 —> 2H2S04 + энергия. |
| Железобактерии | Превращают закисное железо в окисное. Образованная гидроокись оседает и образует так называемую болотную железную руду: 4FeC03 + 02 + 6Н20 -» 4Fe(OH)3 + 4С02 + энергия. В результате жизнедеятельности бактерий в древние геологические периоды образовались залежи железных и марганцевых руд. |
Дата добавления: 2015-12-19; просмотров: 33; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!