Проведение электрического импульса по нервному волокну
Одной из важнейших функций нервной системы – передача нервных импульсов по волокнам от одной клетки к другой. На рисунке 8-6 представлена схема строения нервной клетки. Различают тело нейрона – сому и систему нервных отростков. В клетке имеется большое количество коротких отростков – дендритов, число которых может достигать нескольких тысяч.
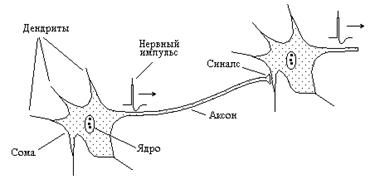
Рис. 8-6
Кроме того, имеются длинные волокна - аксоны. Длина аксонов достаточно велика и у крупных животных может достигать нескольких метров. По дендритам нервные импульсы в нормальных условиях распространяются к телу клетки, по аксонам электрическое возбуждение поступает к эффекторам – тем клеткам, которыми управляет нервная система (мышечным, железистым или к другим нейронам). В последнем случае между аксоном и нейроном образуется синапс, включающий пресинаптическую мембрану аксона, постсинаптическую мембрану клетки и синаптическую щель.
Рассмотрим случай, когда на мембрану тела клетки действует пороговый раздражитель. Этим раздражителем может быть электрический ток, а также медиатор (посредник) – химическое вещество, выделяющееся в синаптическую щель из пресинаптической мембраны. До раздражения нейрона на мембране тела и аксона существует потенциал покоя - цитоплазма клетки (аксоплазма) имеет отрицательный потенциал по отношению к окружающей среде (рис. 8-7 а). При воздействии раздражителя пороговой силы R клетка возбуждается и происходит перезарядка мембраны (генерация потенциала действия) - цитоплазма заряжается положительно. В этом случае на внешней и внутренней поверхности образуются области с различными потенциалами (см. рис. 8-7b). Поскольку в цитоплазме содержатся положительные и отрицательные ионы, возникает локальный электрический ток i. В результате протекания этого тока на участке мембраны аксона, расположенного рядом с телом клетки происходит снижение трансмембранной разности потенциалов. Действительно, на внешней мембране за счет локальных токов притекают отрицательные, а к внутренней поверхности - положительные заряды. Эти заряды нейтрализуют соответствующие потенциалы, и поэтому результирующая разность потенциалов уменьшается. Когда снижение напряжения достигает некоторого критического значения, происходит генерация потенциала действия на участке мембраны аксона, прилежащего к телу клетки. За это время оканчивается цикл возбуждения мембраны клетки и наступает фаза гиперполяризации.
|
|
|
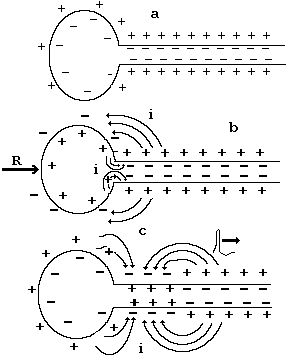
Рис. 8-7
Таким образом, за рассмотренный промежуток времени произошло возбуждение мембраны клетки, возвращение ее в исходное состояние и возбуждение начального участка мембраны аксона.
|
|
|
В данном случае на внешней и внутренней поверхностях мембраны опять возникают области с различными потенциалами, снова протекают локальные токи, снижается мембранный потенциал на последующем участке аксона и генерируется новый потенциал действия. Описанный механизм обеспечивает непрерывное распространение электрического импульса вдоль оси аксона от тела клетки на периферию (см. рис. 8-7с). В нормальных условиях электрический импульс распространяется по аксону от тела клетки. Однако, как следует из рисунка, локальные токи протекают так, что они должны снижать разность потенциалов на участках мембраны, которые уже находились в возбужденном состоянии. Почему же при этом не возникает распространение электрического импульса в противоположном направлении?
Такому аномальному распространению нервного импульса препятствует как, минимум, два условия. Во-первых, на мембране, возвратившейся в исходное состояние, развивается фаза гиперполяризации (повышенного потенциала покоя), и поэтому локального тока оказывается недостаточно, чтобы понизить потенциал покоя докритического значения. Во-вторых, после возбуждения нервная клетка на некоторое время t1 полностью теряет способность к возбуждению. Этот период получил название фазы абсолютной рефрактерности: возбудимость В равна нулю (рис. 8-8).
|
|
|
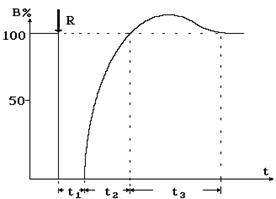
Рис. 8-8
Впоследствии возбудимость постепенно восстанавливается, однако на протяжении времени t2 возбуждение может быть вызвано только такими раздражителями, сила которых превышает пороговую. Промежуток времени t2 называется фазой относительной рефрактерности (относительной невозбудимости). Таким образом, в основе механизма распространения электрического импульса по нервному волокну лежит последовательная деполяризация мембраны аксона в результате протекания локальных электрических токов.
Принципиальное значение для жизнедеятельности всех организмов имеет скорость распространения нервных импульсов. Очевидно, скорость получения, переработки информации и длительность выработки управляющих решений зависит от этого показателя: чем выше скорость проведения, тем быстрее перерабатывается информация. Как показывает теория, скорость распространения электрических импульсов по нервным волокнам зависит от диаметра волокна и электрических свойств мембраны. Чем больше диаметр аксона, тем быстрее распространяется электрический импульс. Именно поэтому у некоторых низших животных в процессе эволюции были использованы нервные волокна с очень большим диаметром. Так, например, у гигантского кальмара диаметр аксона достигает 1 мм. Однако этот путь увеличения скорости оказался неэффективным. Поскольку пропорционально квадрату диаметра возрастает площадь поверхности мембраны, для поддержания неравновесных концентраций требуется значительно больше ионных насосов и соответственно энергетических затрат. Поэтому у высших животных в ходе эволюции было найдено новое решение – изменение электрических свойств мембраны аксона нейрона.
|
|
|
На рисунке 8-9 представлена схема строения нервной клетки, у которой аксон снаружи покрыт цилиндрическим слоем жироподобного вещества – миелина.

Рис. 8-9
Такие волокна получили название миелинизированных или мякотных. Так как миелин относится к жироподобным веществам, он обладает очень высоким электрическим сопротивлением и не пропускает ионы. Миелиновая оболочка прерывается каждые 1-2 мм по длине аксона перехватами Ранвье, имеющими протяженность 1 мкм. В области перехватов мембрана аксона контактирует с окружающей средой. Поэтому разность потенциалов возникает на мембране тела клетки и в перехватах Ранвье (рис. 8-9а). Если на клетку действует раздражитель R пороговой силы, происходит возбуждение мембраны и генерируется потенциал действия (рис. 8-9b). Мембрана тела клетки перезаряжается так, что на внешней поверхности образуется положительный потенциал. При этом, как и в немиелинизированных волокнах, возникают локальные токи i, протекающие между перехватами Ранвье и соответствующими участками мембраны клетки. Эти токи снижают мембранный потенциал в перехвате аксона, и когда он достигается критического значения, в этом участке генерируется новый потенциал действия (рис. 8-9с). Между возбужденным участком аксона и последующим перехватом опять возникают локальные электрические токи i, и процесс повторяется. Таким образом осуществляется распространение нервного импульса между перехватами Ранвье аксона. В отличие от немиелинизированных волокон, в данном случае потенциал действия перескакивает через изолированные миелином участки мембраны на значительно большие расстояния, чем в немиелинизированных волокнах. Именно поэтому скорость проведения электрических импульсов в миелинизированных волокнах существенно больше. Так, например, в немиелинизированных волокнах скорость проведения составляет 0,7-2,5 м/с, а в миелинизированных волокнах такого же диаметра – 10-100 м/с. Описанный механизм проведения электрического импульса в миелинизированных волокнах получил название сальтаторного. Как отмечалось выше, повышение скорости проведения нервных импульсов, достигается в результате изменения электрических свойств (проводимости) мембраны нервного волокна.
Дата добавления: 2016-01-05; просмотров: 46; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!