ДЕРМО-ЭПИДЕРМАЛЬНОЕ СОЕДИНЕНИЕ
АНАТОМИЯ И ГИСТОЛОГИЯ КОЖИ
Кожа представляет собой крупнейший целостный многофункциональный орган, взаимосвязанный со всеми другими органами и системами организма. Непосредственно соприкасаясь с внешней средой, он выполняет барьерно-защитную функцию. Общая площадь поверхности кожи взрослого человека составляет 1,5-1,8 м2, а ее масса вместе с подкожной жировой клетчаткой достигает 17 % массы тела. В области носа, рта, мочеполовых органов и ануса кожа непосредственно переходит в слизистые оболочки. На поверхности кожи наблюдается сложный рисунок в виде треугольных и ромбических полей, сформированный многочисленными бороздками. Более грубые бороздки образуют складки в области ладоней, подошв, мошонки, а также морщины на лице. На ладонных и подошвенных поверхностях пальцев бороздки расположены параллельно и демонстрируют причудливые фигуры, индивидуальные для каждого человека. Эта их особенность используется в криминалистике для идентификации личности (дактилоскопия). Цвет кожи имеет матовый оттенок и обусловлен окраской основных ее тканей, просвечиванием кровеносных сосудов и содержанием пигмента меланина. В зависимости от соотношения этих компонентов у лиц различных рас он варьирует от светло-розового до темно-коричневого. Большая часть кожи покрыта волосами. Исключение составляют ладони, подошвы, боковые, ладонные и подошвенные поверхности пальцев, а также слизистые оболочки губ, ануса и мочеполовых органов. Также на поверхности кожи отмечаются едва заметные отверстия - поры сальных и потовых желез, расположенных в глубине органа. Сальные железы
|
|
|

Рис. 1. Общее строение кожи
отсутствуют в области ладоней и подошв, а потовые - в области слизистой оболочки мочеполовых органов и кожи губ. На тыльных поверхностях дистальных отделов фаланг пальцев кистей и стоп расположены ногти.
Кожа состоит из трех компонентов, или отделов (рис. 1):
• эпидермиса (epidermis);
• дермы (dermis);
• подкожной жировой клетчатки (subcutis), или гиподермы (hypodermis).
Эпидемис является эпителиальной частью кожи, а дерма и гиподерма - соединительнотканной. Толщина кожи (без подкожной жировой клетчатки) колеблется от 0,8 мм в области век до 4-4,5 мм в области ладоней и подошв. Толщина гиподермы варьирует еще более - от полного ее отсутствия на отдельных участках, до 4-6 см в области живота у полных людей. Пограничная зона между эпидермисом и дермой имеет вид волнистой линии вследствие наличия в дерме выростов - сосочков, обусловливающих образование на поверхности кожи гребешков и борозд, формирующих кожный рисунок. В соединительнотканной части кожи (дерме и гиподерме) располагаются нервы, кровеносные и лимфатические сосуды, мышцы. Кроме того, кожа имеет собственные придаточные образования, к которым относятся волосы, сальные и потовые железы, а также ногти. Несмотря на внешнюю простоту, кожа отличается сложным строением. Эпителиальные структуры (эпидермис, волосы, сальные и потовые железы, ногти) развиваются из эктодермы, кожные нервы - из нейроэктодермы, а соединительнотканные волокни-
|
|
|
стые структуры, кровеносные и лимфатические сосуды, жировая клетчатка и мышцы имеют мезодермальное происхождение. Все компоненты кожи находятся в морфофункциональном единстве, обеспечивая множество ее физиологических функций.
ЭПИДЕРМИС
Эпидермис (epidermis) представляет собой многослойный плоский ороговевающий эпителий, обладающий выраженной регенераторной способностью. Толщина его варирует от 0,04 мм в области век до 1,6 мм в области ладоней и подошв. Он расположен на базальной мембране, отделяющей его от дермы, и состоит из клеток - эпидермоцитов. Выделяют пять его слоев:
1) базальный (stratum basale);
2) шиповатый (stratum spinosum);
3) зернистый (stratum granulosum);
4) блестящий (stratum lucidum);
5) роговой (stratum corneum).
Базальный слой располагается непосредственно на базаль-ной мембране и состоит из одного ряда призматических клеток, располагающихся перпендикулярно кожной поверхности. Над базальным слоем расположен шиповатый слой, состоящий из 3-8 рядов клеток многоугольной формы, отростки которых напоминают шипы растений. Мальпигиевым, или ростковым, слоем эпидермиса называется структурная единица, объединяющая базальный и шиповатый слои. Зернистый слой, расположенный над шиповатым, представляет собой 2 ряда (на ладонях и подошвах - 3-4) уплощенно-ромбических клеток. Блестящий слой наблюдается только на ладонях и подошвах. Он выглядит как бесцветная полоска, состоящая из вытянутых безъядерных клеток, расположенных над зернистым слоем. Поверхностным, соприкасающимся с внешней средой, является роговой слой, представляющий собой черепицеобразно расположенные безъядерные роговые чешуйки (корнеоциты). Толщина рогового слоя варьирует от 9 мкм в области век и мужских половых органов до 0,5 мм в области ладоней и подошв. В своей наружной части корнеоциты менее компактны и легко отделяются, обеспечивая процесс физиологического (нормального) шелушения.
|
|
|
В эпидермисе имеются множественные нервные окончания, а его питание осуществляется лимфой через расположенные между клетками межклеточные канальцы, ориентированные перпендикулярно поверхности кожи. Сосудов в эпидермисе нет.
|
|
|
Состав эпидермоцитов неоднороден и представляет собой следующие виды клеток:
1) кератиноциты;
2) меланоциты;
3) клетки Меркеля;
4) клетки Лангерганса;
5) клетки Гренстейна.
Кроме того, в эпидермисе могут встретиться отдельные лимфоциты, а иногда и тканевые базофилы.
1. Кератиноциты - эпителиальные клетки, расположенные во всех слоях эпидермиса, в каждом из которых они имеют различное строение.
Базальные кератиноциты имеют овально-цилиндрическую форму. Их ядра богаты хроматином, а контуры, равно как и плазмалемма, имеют ровные очертания. В цитоплазме базаль-ных клеток имеются фибриллярные образования в виде нежных тонофиламентов (толщиной 3-5 нм) и отдельных пучков - то-нофибрилл, состоящих из белка α-кератина. Их цитоплазма содержит большое количество рибосом и митохондрий, меланосо-мы (гранулы пигмента - меланина, поступающего из меланоци-тов) и лизосомы, осуществляющие фагоцитарную функцию. Между собой они соединяются с помощью десмосом, а к базаль-ной мембране прикреплены полудесмосомами. Десмосомы образованы двумя плотными белковыми образованиями, расположенными друг напротив друга за плазмалеммами соседних клеток.
В состав этих образований входят белки десмоплакин I и II типа, энвоплакин, периплакин и плакоглобин. С ними связываются подходящие к ним пучки внутриклеточных кератиновых тонофиламентов. Связывающие тонкие тонофиламенты, состоящие из белков десмоглеинов и десмоколинов, пронизывают плазмолемму и межклеточное пространство шириной 25-30 нм, и зигзагообразно соединяясь со встречными тонофиламентами соседней клетки, переходят в ее плазмолемму и десмосому. Полудесмосомы представляют собой образования, напоминающие половинки десмосом, прикрепленных к базальной мембране. Базальные кератиноциты осуществляют начало синтеза предшественника кератина - прекератина, который далее превращается в фибриллярный кератин, нити которого скручиваются, формируя кератиновые тонофиламенты. Тонофиламенты образуют каркас базальных ке-ратиноцитов, их десмосом и полудесмосом. Далее базальные филамен-ты постепенно трансформируются в тонофибриллы, уже различимые в световом микроскопе.
Базальные кератиноциты обладают митотической активностью и имеют две субпопуляции клеток. Одна из них постоянно
пролиферирует, в результате чего более молодые клетки остаются на некоторое время на базальной мембране, а более старые "выталкиваются" по направлению к поверхности кожи. В процессе движения они проходят все стадии дифференцировки (шиповатые клетки, зернистые клетки) и превращаются в кор-неоциты. Вторая субпопуляция клеток является резервной и находится в состоянии покоя.
Шиповатые кератиноциты имеют неправильную многоугольную форму. Ядра их округлые, с четко очерченной ядерной мембраной и хорошо просматривающимся ядрышком. В цитоплазме шиповатых клеток имеется большое количество фибриллярных образований, представляющих собой многочисленные пучки различных диаметров и конфигураций, рибосом, обеспечивающих усиление синтеза прекератина, а также сохраняются меланосомы. Десмосомы шиповатых клеток (узелки Биццоце-ро) более крупные - до 300-700 нм. Сцепление клеток шиповатого слоя осуществляется как за счет крупных десмосом, так и за счет проникновения цитоплазматических отростков одних клеток в углубления других (по типу "застегнутой молнии").
Зернистые кератиноциты имеют вытянутую параллельно поверхности кожи уплощенно-ромбическую форму. Ядра клеток обладают плеоморфизмом (округлые, овальные, вытянутые, лишенные ядрышка, подвергшиеся кариорексису и т. д.), а плаз-малемма на отдельных участках имеет зазубренные контуры. В цитоплазме встречается множество гранул кератогиалина, хорошо окрашенных и имеющих вид зерен с диаметром в несколько микрометров и, соответственно, хорошо различимых в световом микроскопе. Ни кератина, ни гиалина эти гранулы не содержат. Вместе с тонофибриллами они образуют тонофибриллярно-кера-тогиалиновые комплексы, обеспечивающие синтез филагрина - основного белка гранул.
Филагрин вместе с филаментами обеспечивает правильное расположение и сцепление корнеоцитов. Зернистые кератиноциты продолжают синтез прекератина, с уже большей молекулярной массой, чем у прекератина, синтезируемого базальными и шиповатыми клетками. Кроме того, в периферических отделах цитоплазмы зернистых клеток имеются кератиносомы (видоизмененные лизосомы или гранулы Од-ланда). Они содержат набор веществ (гидролитические ферменты, гли-колипиды, гликопротеиды, стерины), обеспечивающих образование межклеточного цементирующего гидрофобного вещества, выполняющего барьерную функцию рогового слоя эпидермиса.
Корнеоциты (ранее называвшиеся роговыми чешуйками) представляют собой безъядерные удлиненные клетки, располо-
женные черепицеобразно параллельно поверхности кожи. Они имеют длину до 10-12 мкм и толщину от 0,1 до 1 мкм. Сцепляются корнеоциты как за счет взаимопроникающих выростов плотной оболочки, так и остатков ороговевших десмосом. Кор-неоцит заполнен аморфным веществом, пронизанным керати-новыми фибриллами. В нижних рядах отдельных клеток сохраняется значительное количество гранул кератогиалина, что микроскопически соответствует блестящему слою эпидермиса. В небольших количествах роговые клетки содержат зрелый кератин.
2. Меланоциты - отросчатые клетки нейроглиального происхождения, расположенные в базальном слое эпидермиса и отвечающие за синтез меланина. Меланин представляет собой пигмент, защищающий кожу от вредного воздействия ультрафиолетовых лучей и придающий ей коричневый оттенок. Мела-ноциты имеют ядра с неровными очертаниями мембраны и многочисленные отростки, в цитоплазме которых содержатся множественные меланосомы - структуры, осуществляющие биосинтез меланина. Отростки клеток при помощи десмосом и полудесмосом соединяют меланоцит с несколькими соседними кератиноцитами, меланоцитами и базальной мембраной.
Меланин синтезируется из тирозина под воздействием каталитических ферментов - тирозиназы и ДОФА-оксидазы и посредством экзо-цитоза передается в межклеточное пространство, откуда и фагоцитируется кератиноцитами. Меланогенез с помощью меланоцитстимулирую-щего гормона контролируется гипофизом. В его процессе также участвуют ионы меди, витамин С, адреналин и норадреналин, гормоны щитовидных и половых желез. Цвет кожи и степень ее защиты от излучения зависят от количества меланина в кератиноцитах, а не от количества самих меланоцитов, синтез меланина в которых увеличивается под воздействием ультрафиолетового облучения.
3. Клетки Меркеля представляют собой осязательные нейро-эндокринные клетки, расположенные в базальном и шиповатом слоях эпидермиса, наружных слоях эпителия волосяного фолликула и некоторых слизистых оболочках. Они несколько крупнее кератиноцитов, имеют округлую или вытянутую форму, содержат удлиненное и слегка сегментированное ядро. В их цитоплазме располагаются филаменты цитокератинов и специфичные для этих клеток нейроэндокринные гранулы диаметром 100-200 нм.
В гранулах содержится комплекс нейропептидов (мет-инкефалин, фактор роста нервов, вазоинтестинальный полипептид, бомбезид, субстанция Р, эндорфины). С кератиноцитами клетки Меркеля соединя-
ются при помощи десмосом, а с нервными окончаниями в зоне своего нижнего полюса они формируют образования, напоминающие синапсы. В области этих образований в цитоплазме концентрируются нейро-эндокринные гранулы, а между плазмолеммой клеток и нервными окончаниями имеется синаптическая щель размером 50 нм. Некоторые клетки Меркеля свободны и не связаны с нервными волокнами.
4. Клетки Лангерганса - дендритические клетки, имеющие моноцитарное происхождение. Они относятся к системе фагоцитирующих мононуклеаров и являются тканевыми макрофагами. Эти клетки располагаются как в базальном, так и в нижних рядах шиповатого слоя эпидермиса. Их ядра имеют сегментированную форму и 1-2 ядрышка среднего размера. Цитоплазма клеток светлая и содержит множество органелл, среди которых встречаются особые структуры - так называемые гранулы Бирбека в форме теннисной ракетки, размерами от 30 нм в ширину до 200 нм в длину. Роль этих гранул полностью не установлена, однако известно, что в них накапливается эпидермальный кейлон, подавляющий пролиферацию кератиноцитов. Клетки Лангерганса имеют многочисленные отростки, с помощью которых соединяются друг с другом и с соседними кератиноцитами, образуя характерную ветвистую сеть. Отростки нижнего слоя клеток проникают в дерму.
Клетки Лангерганса не являются оседлыми и постоянно мигрируют в дерму и лимфатические узлы, выполняя свою макро-фагальную функцию. Они осуществляют процессинг и презентацию антигенов Т-хелперам (как проникающим в эпидермис, так и находящимся в дерме или лимфатических узлах), а также сек-ретируют ряд медиаторов (интерлейкины-1, -4, γ-интерферон, фактор некроза опухолей и др.), регулирующих пролиферацию и дифференцировку Т-лимфоцитов и других иммунокомпетент-ных клеток. Количество клеток Лангерганса составляет около 3-4 % всех эпидермоцитов.
5. Клетки Гренстейна представляют собой клетки, относящиеся к популяции дендритических клеток. Морфологически они весьма напоминают клетки Лангерганса, образуя похожую ветвистую сеть, но отличаются от них главным образом отсутствием гранул Бирбека и некоторых макрофагальных мембранных антигенов (HLA-DR). В то же время клетки Гренстейна экс-прессируют молекулы, весьма напоминающие антиген - маркер тимоцитов, таким образом свидетельствуя, что они иммуноло-гически отличаются от клеток Лангерганса. Функция этих клеток окончательно еще не установлена.
ДЕРМО-ЭПИДЕРМАЛЬНОЕ СОЕДИНЕНИЕ
Дермо-эпидермальное соединение (рис. 2) представляет собой сложную специализированную межклеточную структуру (базальную мембрану) толщиной 0,7-1,0 мкм, с помощью которой эпидермис прикрепляется к дерме. Эта структура состоит из следующих четырех компонентов:
1) клеточных мембран и полудесмосом базальных эпидер-моцитов;
2) светлой пластинки (lamina lucida);
3) плотной пластинки (lamina densa);
4) фиброретикулярной пластинки.
Полудесмосомы являются участками утолщения плазмолеммы ба-зальных кератиноцитов, соединенных с внутриклеточными кератино-выми тонофиламентами. Эти утолщения состоят из плакиновых белков, формирующих так называемый "антиген буллезного пемфигоида I типа" (BP 230). Кроме того, они содержат также плектин, от которого в сторону светлой пластинки отходят пронизывающие плазмолемму
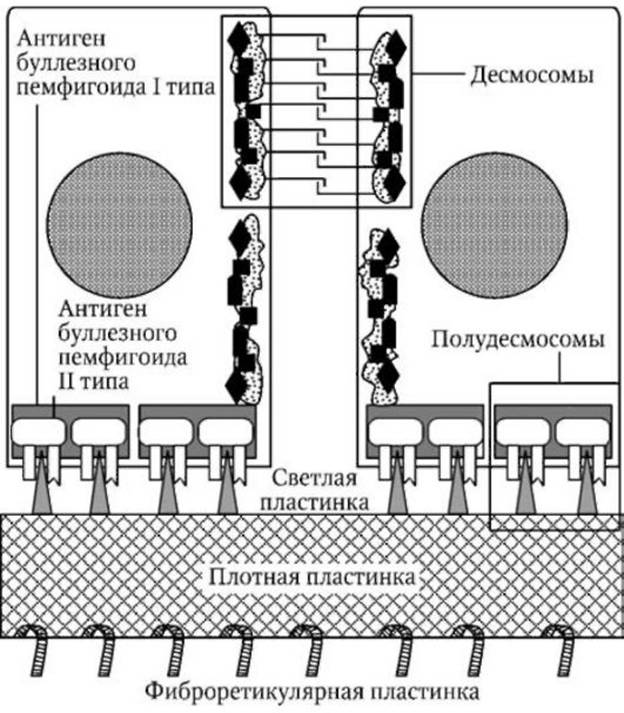
Рис. 2. Строение базального слоя и базальной мембраны эпидермиса
белковые тонофиламенты, состоящие из "антигена буллезного пемфи-гоида II типа" (BP 180) и интегрина.
Светлая пластинка (толщиной 30-40 нм) представляется преимущественно аморфной, однако в центральной части она содержит плотную линию, вертикально соединенную с внеклеточными тонофиламен-тами базальных полудесмосом. Светлая пластинка содержит ламинин, нидоген, энтактин, фибронектин и коллаген IV типа.
Плотная пластинка (толщиной 30-60 нм) состоит из волокнистого материала, скрепленного аморфным веществом. В ней содержится коллаген IV и V типа, протеогепарансульфат и ламинин.
Фиброретикулярная пластинка представляет собой субэпидермаль-ное сплетение соединительнотканных волокон, соединяющее дерму с базальной мембраной. Она образована пучкообразными крепящими (якорными) фибриллами, расположенными вертикально и соединенными верхними концами с плотной пластинкой, а также микрофибриллами эластических волокон и одиночными коллагеновыми волокнами.
Дермо-эпидермальное соединение выполняет как опорную функцию, так и функцию проникновения и транспортировки физиологических веществ и клеток из дермы в эпидермис и обратно.
ДЕРМА
Дерма (dermis) состоит из:
1) волокнистой соединительной ткани;
2) основного (аморфного) вещества;
3) клеточных элементов.
Волокнистая соединительная ткань и основное вещество объединяются в так называемое межклеточное вещество. Дерма в 15-40 раз толще эпидермиса и, помимо основных своих компонентов, содержит придатки кожи (волосы, сальные и потовые железы), кровеносные и лимфатические сосуды, мышцы и нервы.
Анатомически дерма разделяется на сосочковую и сетчатую (ретикулярную) части. Верхний ее отдел, представленный сосочками, вдающимися в эпидермис, относится к сосочковой дерме. Нижняя ее часть, сверху ограниченная концевыми отделами эпидермальных отростков, а снизу без видимой границы переходящая в гиподерму, - к ретикулярной. Кроме того, выделяют еще и периаднексальную дерму, подразумевая под этим понятием тот ее отдел, который непосредственно окружает придатки кожи. Сосочковая дерма вместе с периаднексальной объединяются в так называемую адвентициальную дерму, представляющую собой морфофункциональную единицу.
Волокнистая соединительная ткань состоит из коллаге-новых, эластических и ретикулиновых волокон.
1. Коллагеновые волокна сгруппированы в пучки, образованные фибриллярным белком - коллагеном, и обеспечивают функцию механического каркаса дермы. В сосочковом слое эти пучки мелкие и ориентированы хаотично. В сетчатом слое они более грубые и располагаются параллельно поверхности кожи.
Каждое коллагеновое волокно состоит из коллагеновых фибрилл (толщиной 40-50 нм), упакованных в определенном порядке и склеенных цементирующим веществом. По полипептидному строению в со-сочковом слое в основном располагается коллаген III типа, а в сетчатом - I типа. В состав их аминокислотных цепей входят пролин, окси-пролин, глицин, лизин и оксилизин. Часть лизина и пролина остается в коллагене в гидроксилированном виде, участвуя в образовании химических связей, обеспечивающих особую прочность коллагена.
2. Эластические волокна обеспечивают функцию возврата кожи в первоначальное состояние после ее растяжения. Они состоят из фибриллярной (гликопротеидной) части и аморфной, представляющей собой белок - эластин.
Соотношение этих компонентов в волокнах, расположенных в разных отделах дермы, неодинаково. В сосочковой части в волокнах преобладает гликопротеидный компонент, обеспечивая им необходимую прочность для участия в формировании базальной мембраны эпидермиса. В сетчатой части расположены "зрелые" эластические волокна, максимально насыщенные эластином (до 80-95 %), отвечающим за их эластические свойства. В состав эластина входят полипептидные цепи, содержащие преимущественно аланин, пролин, десмозин, глицин и валин.
3. Ретикулиновые волокна наиболее распостранены в периад-нексальной дерме и располагаются преимущественно вокруг сосудов, волосяных фолликулов, сальных и потовых желез. В связи с их свойствами окрашиваться нитратом серебра и не окрашиваться обычными методами, они также называются аргирофиль-ными. Ретикулиновые волокна состоят как из гликопротеидно-го, так и фибриллярного компонента, представляющего собой коллаген III типа. Напоминая по своему строению коллагено-вые, эти волокна выполняют функцию механического каркаса для сосудов и придатков кожи.
Основное вещество представляет собой комплекс гликоза-миногликанов, гликопротеидов и протеогликанов. Также оно содержит жиры, неорганические вещества и воду.
1. Гликозаминогликанами дермы являются хондроитин-(4,6)-сульфат, гепарин, гиалуроновая кислота и дерматансульфат.
Их молекулы представляют собой длинные цепи повторяющихся сахаров. Благодаря неплотной пространственной упаковке полисахаридов, гликозаминогликаны обладают способностью поглощать и удерживать большое количество воды, создавая необходимые условия как для клеточных реакций, так и для биохимических процессов. Гиалуроновая кислота не образует комплексов с белками, быстро повреждается гиалуронидазой, осуществляя еще большую проницаемость основного вещества.
2. Гликопротеиды представляют собой соединения длинных полипептидных цепей с короткими разветвленными цепями са-харидов. К наиболее важным из них относится фибронектин, играющий значительную роль в адгезии и кооперации клеток соединительной ткани, образовании коллагеновых волокон и развитии иммунных реакций.
3. Молекулы протеогликанов состоят из длинных белковых цепей, по бокам соединенных с большим количеством нераз-ветвленных цепей гликозаминогликанов. Они обеспечивают гидратацию, прочность основного вещества, его способность деформироваться и восстанавливаться.
Клеточные элементы дермы представлены фибробластами, тучными клетками, гистиоцитами, а также, в незначительном количестве, лимфоцитами и эозинофильными лейкоцитами.
1. Фибробласты - неоднородная популяция клеток, обеспечивающая синтез волокнистой соединительной ткани и основного вещества. К фибробластам относятся слабодифференциро-ванные фибробласты, дифференцированные фибробласты, фиброциты и фиброкласты. Они приступают к дифференцировке, образуясь из предшественников - перицитов - клеток, входящих в состав стенок кровеносных сосудов.
Слабодифференцированные фибробласты имеют звездчатую форму, округлое ядро и базофильную цитоплазму со слаборазвитой эндоплаз-матической сетью. Дифференцированные фибробласты имеют 12-20 мкм в диаметре, большие светлые ядра с крупными ядрышками, слегка ба-зофильную цитоплазму, в которой хорошо развиты клеточные орга-неллы. Среди последних особенно заметны характерные фибриллы. Дифференцированные фибробласты постепенно переходят в фиброциты - функционально неактивную субпопуляцию фибробластов, имеющих веретенообразную форму, вытянутое гиперхромное ядро и частично деградированные органеллы. На рибосомах фибробластов осуществляется синтез начальных полипептидных цепей коллагена, которые далее, после гидроксилирования и гликозилирования, сшиваются ди-сульфидными мостиками в трехцепочечную спираль тропоколлагена. После завершения процесса создания молекулы тропоколлагена фиб-
робласты его секретируют и дальнейший синтез зрелого коллагена осуществляется уже внеклеточно. Менее изучен процесс синтеза эластических и аргирофильных волокон, осуществляемый другими субпопуляциями фибробластов. Обновление межклеточного вещества происходит за счет его разрушения другой субпопуляцией клеток - фиброкластами, выполняющими фагоцитарную функцию и гидролиз соединительной ткани с помощью своих лизосом.
2. Тучные клетки (лаброциты, мастоциты) - клетки, являющиеся потомками стволовой клетки крови и тканевыми аналогами базофильных лейкоцитов крови. Лаброциты характеризуются плеоморфизмом, отличаясь друг от друга как размерами, так и очертаниями. Ядра клеток крупные, округлые или овальные. Характерная особенность этих клеток - наличие в их цитоплазме специфических гранул, содержащих биологически активные вещества: гистамин, гепарин, серотонин, хондроитин-сульфат, гиалуроновую кислоту и др. Содержимое этих гранул оказывает регулирующее влияние на проницаемость сосудов кожи, количество воды в межклеточном веществе дермы, пролиферацию и дифференцировку различных клеток, синтез гликоз-аминогликанов.
3. Гистиоциты представляют собой дермальные макрофаги и относятся к системе фагоцитирующих мононуклеаров. Они имеют очертания неправильной формы, образованные многочисленными отростками. Ядра клеток крупные, темные, бобовидной формы с отдельными инвагинациями. Цитоплазма ва-куолизирована и имеет хорошо развитые многочисленные лизо-сомы, которые с помощью своих лизосомальных ферментов осуществляют главную функцию макрофагов - фагоцитоз. Если же захваченные чужеродные вещества не могут быть переработаны окончательно, макрофаги осуществляют их процессинг, переводя антигены веществ в высокоиммунную форму. Далее макрофаги, вместе с собственным антигеном, экспрессируют переработанный продукт на своей поверхности для последующей кооперации его с лимфоцитами. Кроме того, макрофаги секре-тируют различные медиаторы межклеточного взаимодействия (интерлейкин-1, γ-интерферон, фактор некроза опухолей и др.), осуществляющие регуляцию пролиферации и дифференцировки иммунокомпетентных клеток и фибробластов. В цитоплазме некоторых гистиоцитов обнаруживаются гранулы Бирбека, свидетельствующие о том, что эти макрофаги являются мигрирующими из эпидермиса и обратно в эпидермис клетками Лангерганса.
Дата добавления: 2021-01-20; просмотров: 157; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!