О подклассе Batrachosauria — группе форм, промежуточных между земноводными и пресмыкающимися, 1946 г.
ИЗВЕСТИЯ АКАДЕМИИ НАУК СОЮЗА ССР
Отделение биологич. наук № 6, 1946
(Представлено академиком И. И. Шмальгаузеном)
В песках известного Ишеевского местонахождения (Каменный овраг близ дер. Теникеево Апастовского района Татреспублики) в Среднем Поволжье, относящихся к II зоне верхней перми, раскопками Палеонтологического института Академии Наук СССР были обнаружены обломки костей необыкновенного животного.
В 1934 и 1935 гг. среди множества остатков травоядных и хищных дейноцефалов и нескольких костей стегоцефалов типа Platyops было найдено несколько кусочков костей крыши черепа с необыкновенной «балочной» скульптурой. Микроскопическое исследование этих обломков, предпринятое по моей просьбе А. П. Быстровым, показало значительное отличие их строения от костей крыши черепа типичных стегоцефалов, в частности лабиринтодонтов Benthosuchus и Platyops. В то же время микроструктура найденных обломков ничем существенным не отличалась от таковой у рептилий, точнее от микроструктуры покровных костей черепа котилозавров.
Вместе с обломками черепных костей были найдены разрозненные позвонки различной величины, принадлежавшие разным особям. Строение этих позвонков, обладавших очень широкими и массивными невральными дугами, было настолько типичным и настолько сходным с строением позвонков Seymouria, что я без колебания отнес их к примитивным Cotylosauria (Ефремов, 1940, табл. VI. фиг. 1—4). Характер строения животного по обломкам невиданных ранее черепных костей, разумеется, не поддавался определению. Все же я заключил, что имею дело с обломками parietalia и nasalia сравнительно плоского и широкого черепа, могущего быть черепом стегоцефала, и предложил для загадочной формы условное название Lanthanosuchus qualeni, отнеся ее к группе сеймурий (Ефремов, 1941). Новые раскопки Палеонтологического института в Ишеевском местонахождении в 1939 г., заложенные на северной окраине костеносной линзы, дали ряд остатков стегоцефалов Platyops, Melosaurus и еще двух новых форм лабиринтодонтов, помимо костей дейноцефалов и челюстей Venjukovia. В самой середине вскрытого участка костеносного песка был найден совершенно полный, превосходной сохранности череп необыкновенного животного, который был сразу же отождествлен с ранее найденными фрагментами черепных костей Lanthanosuchus. Кроме черепа, были найдены отдельные мацерированные элементы крыши черепа (quadratojugalia и squamosa), отдельные неполные позвонки и плохо определимый фрагмент нижней челюсти, могущий быть отнесенным к этому же животному, однако, не дающий представления о ее строении.
|
|
|
Таким образом от посткраниального скелета новой находки мы имеем лишь разрозненные позвонки и в сущности можем говорить лишь о строении его черепа, без нижней челюсти. Тем не менее открытие этой необычайной формы представляет большой интерес и имеет важное значение для познания древнейших этапов эволюции рептилий.
|
|
|

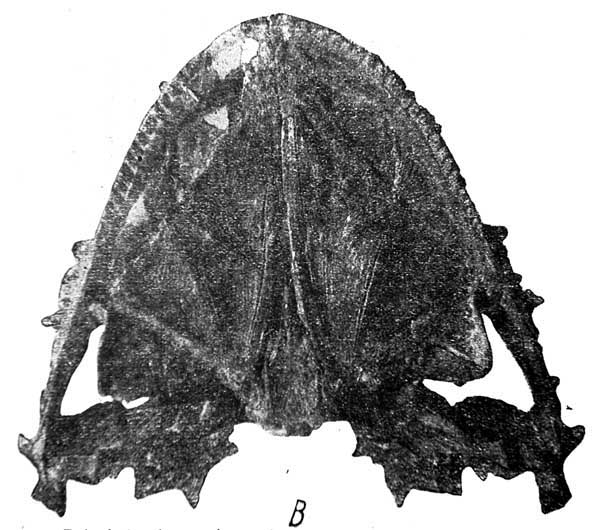
Табл. I. Lanthanosuchus watsoni, gen. et sp. nov. Череп № 271/1. A — череп сверху; B — череп снизу, 4/7 натуральной величины.
Череп № 271/1 (табл. I, А н В) был отпрепарирован препаратором института М. Ф. Лукьяновой так, что ни одна из важнейших деталей его строения не осталась скрытой. В этом отношении череп практически не отличается от любого из черепов современных нам животных в зоологических коллекциях.
А. П. Быстров любезно выполнил ряд рисунков черепа. На этих рисунках ни одна подробность не реконструирована. Каждый рисунок представляет точную ортопроекцию натуры.
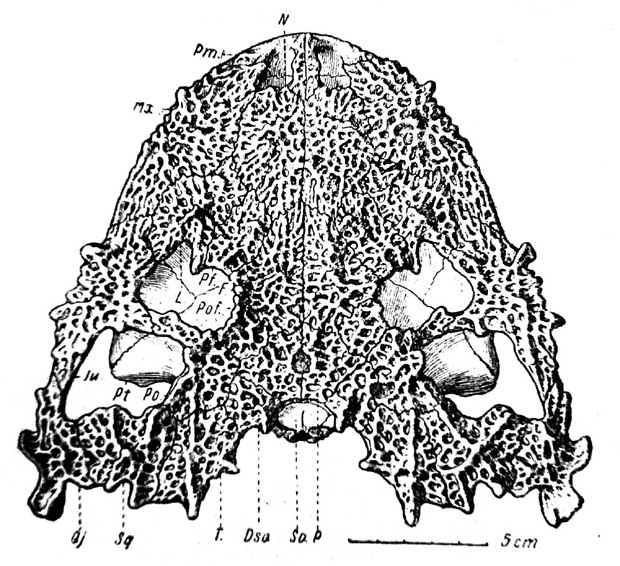
Рис. 1. Lanthanosuchus watsoni, gen. ер. nov. Череп № 271/1, сверху.
Dso — dermosupraoccipitale; F —frontale; Ju — jugale; Mx — maxillare; L — lacrymale; N — nasale; P — parietale; Рm — praemaxillare; Pt — pterygoideum; Pf — praefrontale; Po — postorbitale; Pof — postfrontale; Qj — quadratojugale; So — supraoccipitale; Sq — squamosum; T — tabulare
Дорзальная поверхность черепа (рис. 1). Череп широкий и очень плоский, параболической формы. Длина по средней линии от переднего края морды до заднего конца шва между parietalia в глубине затылочного выреза 140 мм. Ширина между задними отростками quadratojugalia 205 мм.
|
|
|
Ноздри сближены между собою и разделены узкой перегородкой, образованной задними отростками praemaxillaria,. как у всех Cotylosauria и Theromorpha. В дне носовой полости выходит хорошо развитое septomaxillare с вырезкой для назолакримального протока.
Орбиты слегка отодвинуты назад от середины длины черепа и сближены между собою; их медиальные края сильно приподняты, отчего межорбитальная поверхность черепа имеет корытообразный вид. Орбиты имеют круглую форму, осложненную большими подглазничными фонтанелями, как у древнейших эмболомерных амфибий карбона. Слуховые вырезки отсутствуют, затылочный край черепа в середине врезан вперед крутой дугой. По сторонам задние края крыши черепа отвернуты вверх и осложнены отростками и пластинчатыми ребристыми выступами.
В посторбитальном отделе черепа крайне интересно наличие больших височных ям, построенных в смысле ограничения элементами крыши черепа, по типу нижней височной ямы Therapsida.
Кости крыши черепа покрыты необыкновенной для стегоцефалов очень грубой ямчато-бугорчатой скульптурой, осложненной сетью перекрещивающихся балок, валиков и бугорков, похожих на пластинчатые стойки, сверху закругленные.
|
|
|
Praemaxillaria — широкие, поперечно вытянутые, их поверхность почти лишена скульптуры, за исключением задне-верхних отростков, образующих носовую перегородку. Последние несут спереди парные, сливающиеся в один, высокие бугорки. Позади бугорков поверхность отростков скульптирована.
Махillaria — очень большие, в центрах своего окостенения несут большие, торчащие вверх и вбок костные выросты.
Nasa1ia — широкие, слегка суживаются назад, имеют выпуклую в центре поверхность.
Frontalia — треугольной формы, вместе образуют фигуру ромба, латеральные углы которого свободны, закруглены, утолщены и отвернуты вверх, образуя средние части внутренних краев орбит. Посередине передних углов frontalia проходят сильные валикообразные выпуклости.
Praefrontalia — довольно большие, ступенчато изогнутые, соответственно подъему орбитальных краев, далеко разобщенные от postfrontalia.
Laсгуmalia — большие и широкие. Их передние концы достигают лишь половины расстояния между орбитами и ноздрями. Таким образом эти кости являются сравнительно укороченными, как у стегоцефалов. В центре окостенения каждое lacrymale несет высокий костный бугорок. Задний, примыкающий к орбите край lacrymale свободен, закруглен и несет глубокую треугольную вырезку, образующую передний угол суборбитальной ямы или фонтанели. В состав орбитального кольца входит только небольшой участок постеромедиальной оконечности lacrymale, вытянутый в закругленный отросток, разграничивающий орбиту и суборбитальную яму. Вся остальная часть lacrymale находится в пределах суборбитальной ямы.
Jugalia — большие кости сложной конфигурации. Передний край каждого jugale граничит с lacrymale на середине суборбиталъной ямы, передний, углообразно заостренный конец кости вклинивается между lacrymale и maxillare значительно впереди от конца суборбитальной ямы. Внешний свободный край jugale несет два больших костных выроста и несколько мелких.
Внутренние края jugale также свободны, в середине несут широкий отросток, соединяющийся с postorbitale в середине заднего края суборбитальной ямы и составляющий вместе с последним посторбитальную перегородку между орбитой и височной ямой. Задний конец jugale сужен, вытянут и налегает косым швом на quadratojugale в пределах задней трети длины височной ямы.
Postfrontalia — маленькие, субквадратных очертаний, каждое из них составляет примерно половину верхнего края орбиты.
Postorbitalia — велики и серповидно изогнуты. Позади своих латеральных отростков, входящих в состав посторбитальных перегородок, кости продолжаются далеко назад до середины верхнего края височных ям, сочленяясь с squamosa и tabularia позади, с dermosupraoccipitalia медиально(!) и с postfrontalia спереди. Только маленькие внешние углы parietalia соприкасаются с внутренними краями postorbitalia, что представляет собою одну из многих интересных особенностей описываемого черепа.
Parietalia — укорочены и расширены, имеют совместно очертания сплющенного ромба. В середине длины шва между обеими костями располагается очень большое париетальное отверстие с приподнятыми. краями. Как уже упоминалось, parietalia почти целиком оттеснены oт postorbitalia вклиненными сзади костями, которые я определяю пока как dermosupraoccipitalia. Задние края parietalia приподняты и закруглены, к ним наискось снизу причленяется широкое и гладкое supraoccipitale.Эта последняя кость входит в глубокую дугообразную вырезку затылочного края. Таким образом в этом черепе мы встречаемся с необыкновенным обстоятельством, что dermosupraoccipitale не соединяются лруг с другом по средней линии, а широко раздвинуты и разобщены вклиненным между ними supraoccipitale, с которым не контактируют. Поэтому не исключена возможность, что кости определенные условно как dermosupraoccipitalia, на самом деле суть supratemporalia, отодвинутые очень далеко назад и вошедшие в состав затылочного края крыши черепа, как это имеет место у некоторых примитивных котилозавров. Этому последнему определению мешает их непосредственное налегание на paroccipitalia.
Dermosupraoccipitalia (?supratemporalia) вклиниваются очень далеко вперед между postorbitalia и parietalia. Задние свободные края этих костей отвернуты кверху и снабжены симметричными, направленными назад и вверх выступами, в форме поставленных на ребро пластинок.
Tabularia — очень велики и выдвинуты далеко назад; они образуют род костного навеса над затылочной поверхностью черепа. Поверхность костей прогнута соответственно отвороту вверх их задних краев. Последние утолщены и снабжены костными выступами, симметрично расположенными под прямым углом друг к другу на каждой кости: один — внутрь, один — назад. В продолжение каждого выступа по наружной и внутренней поверхностям tabulare проходят длинные валики, как бы костные балки для увеличения прочности.
Squamosa — большие и широкие, имеют такие же балки, скрещивающиеся в виде прямого креста. Затылочный край каждого squamozum несет глубокую вырезку, обрамленную двумя костными отростками. Передний край squamosum в височной яме также имеет тупоугольную вырезку. Squamosa разобщены от jugalia и сочленяются по своему нижнему краю только с quadratojugalia, так как задний угол каждой височной ямы врезан в quadratojugale.
Quadratоjugа1ia — очень большие кости своеобразной формы. Спереди каждое quadratojugale имеет суженный отросток, сочленяющийся с jugale. Верхний край кости сочленен широким швом со squamosum. Задне-внешние свободные стороны quadratojugalia, составляющие внешние углы черепа, снабжены большими симметричными костными выростами в виде толстых лепестков, направленных вперед и назад под прямым углом друг к другу. С внешней стороны поверхность каждого лепестка вогнута (рис. 3, В).
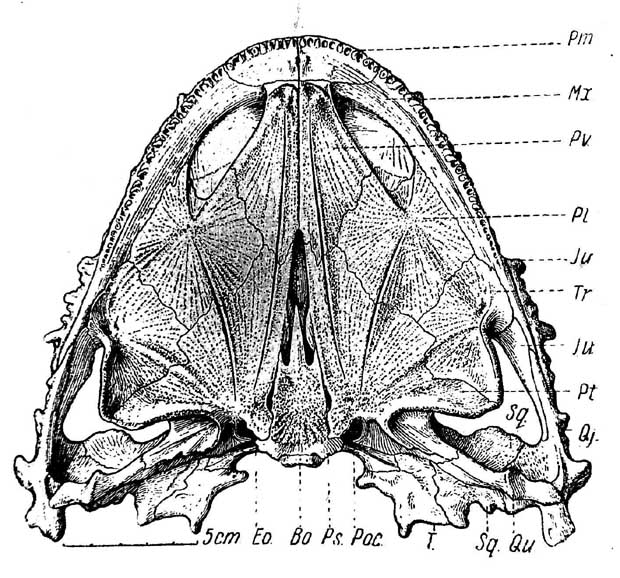
Рис. 2. Lanthanosuchus watsoni, gen. et sp. nov. Череп № 271/1, снизу.
Обозначения как на рис. 1 и еще: Во — basioccipitale; Eo — exoccipitale; Qu — quadratum; Pt — palatinum; Рос — paroccipitale; Ps — parasphenoideum; Pv — praevomer; Tr — transversum; Sq — squamosum.
Вентральная поверхность черепа (рис. 2) характеризуется весьма архаическим типом строения. Хоаны сближены и разделены сравнительно узкой перегородкой, составленной суженными передними концами обоих praevomera. Назад и наружу хоаны переходят в обширные плоские углубления, точнее — вдавленности, овальной формы, постепенно сглаживающиеся кзади. Внутренние стороны этих углублений ограничены клинообразно расходящимися назад краями praevomera, продолженными гребнями на palatina.
Все кости небной поверхности — praevomera, palatina, transversa и pterygoidea — очень большие и широкие, образуют сплошную костную поверхность, как у эмболомерных стегоцефалов. Межптеригоидное отверстие очень узкое и длинное. Только самая передняя оконечность его ограничена praevomera, так как узкие отростки обоих птеригоидов проходят вперед с внутренних сторон задних расходящихся отростков praevomera.
Интересной особенностью является развитие мощных поперечных птеригоидных флангов, обладающих тем же характером строения, как и у тероморфных рептилий.
Широкое, веерообразно развернутое transversum сочленяется с jugale лишь своей передней половиной. Задняя половина латерального края transversum свободна, снабжена утолщенным ребром и приподнята над небной поверхностью, прикрепляясь массивной валикообразной ножкой к jugale. Эта часть transversum образует наружную сторону птеригоидного фланга. Весь задний, сильно завернутый вниз край фланга образован за счет pterygoideum.
Квадратная ветвь птеригоида короткая и сочленяется с длинным передним отростком quadratum на первой трети расстояния от птеригоидного фланга до заднего края инфратемпоральной ямы.
Jugaliа на вентральной поверхности черепа заходят далеко назад, налегают на внутренние стороны quadratojugalia и оканчиваются у задних углов височных ям.
Quadrata образуют широкие катушкообразные поверхности челюстных суставов, опираясь на скошенные нижние выступы quadratojugalia. Каждое quadratum налегает на quadratojugale спереди, так, что нижний край последнего доходит почти до суставной поверхности и вся суставная катушка лежит целиком под quadratojugale.
С внутренней стороны quadratum несет большой округленный бугорок, образующий внутреннюю часть суставной поверхности. Выше этого бугорка quadratum дает длинный отросток, снизу валикообразно закругленный и направленный внутрь, вперед и вверх, до соединения с квадратной ветвью птеригоида. Верхний край этого отростка соединяется вначале с quadratojugale, а затем на всем своем протяжении с squamosum, что хорошо видно на затылочной поверхности (рис. 4).
Parasphenoideum — большой, имеет очертания очень высокой трапеции. Задний край corpus parasphenoidei расширен и утоньшен, налегает снизу на хорошо окостеневшее basioccipitale, узкая полоска которого выступает на вентральной поверхности в неглубокой вырезке средней части заднего края парасфеноида. Постеролатеральные углы парасфеноида выступают назад и наружу свободными пластинками.
Processus cultriformis парасфеноида очень короткий, мечевидно заострен и достигает только половины межптеригоидного отверстия.
Сочленение птеригоидов с basis cranii осуществляется с помощью парасфеноида; базиптеригоидные отростки basisphenoideum, равно как и сам basisphenoideum, не окостеневали. Базальные части птеригоидов образуют сравнительно узкие отростки, сочленяющиеся с телом парасфеноида в его задней половине. Соединение не представляет собою зубчатого шва, а скорее является плотным прилеганием отростков птеригоида, входящих в специальные углубления тела парасфеноида.
Впереди сочленения птеригоидов с парасфеноидом небольшие вырезки во внутренних краях птеригоидов образуют узкие просветы — отверстия для сонных артерий, приходящиеся против середины corpus parasphenoidei. Еще дальше вперед птеригоиды снова вступают в тесное соприкосновение с парасфеноидом. Это второе соединение усилено маленькими отростками в передней части тела парасфеноида, выступающими вперед параллельно processus cultriformis, так, что вся передняя часть парасфеноида похожа на трезубец с удлиненным средним зубцом. Эти заостренные боковые отростки достигают основания восходящих гребешков птеригоидов, тонкими пластинками поднимающихся на дорзалъной стороне костей.
В просветах между средней частью corpus parasphenoidei и птеригоидами, повидимому, проходили сонные артерии (art. carotis interna).
В глубине межптеригоидного отверстия видно костное образование, форма которого не может быть установлена с точностью, так как именно в этом небольшом участке имеется местная посмертная деформация — вдавление с переломом конца processus cultriformis. Этот костный элемент по местоположению соответствует сфенэтмоиду, но несомненно состоит из двух парных частей и, повидимому, является орбитосфеноидом (orbitosphenoideum) — элементом, характерным для рептилий.
Озубление крайне своеобразно (рис. 3, А). Зубы сравнительно тонкие и длинные, загнутые внутрь и назад, расположены в единственный ряд на praemaxillaria и maxillaria. Дополнительные зубные ряды отсутствуют, зато почти вся небная поверхность покрыта резкой шагренью, ориентированной рядами, радиально расходящимися из центров окостенения каждой кости неба. Свободны от шагрени только краевые элементы черепа — praemaxillaria, maxillaria, jugalia и т. д.
Наиболее крупные шагреневые зубы располагаются на дугообразных выступах в конце каждого praevomer и на утолщениях нижнего края птеригоидного фланга (как на pterygoideum, так и на transversum). Некоторое укрупнение шагрени отмечается на свободных внешних краях praevomer, в базальных частях птеригоидов и в задней половине corpus parasphenoidei. Такое же укрупнение наблюдается на невысоком гребне, пересекающем наискось pterygoideum и transversum, у основания поперечного фланга.
Картина распределения шагрени осложняется четырьмя глубокими бороздами, располагающимися симметрично по две с каждой стороны. Одна, более длинная, бороздка проходит от переднего конца praevomer назад почти параллельно срединному шву до базальной части птеригоида. Вторая бороздка идет от базальной части птеригоида к центру окостенения palatinum. Вместе с упомянутым выше гребнем в основании фланга бороздки образуют три радиально расходящихся луча, соединяющих, что весьма существенно, центр окостенения pterygoideum с центром окостенения transversum, palatinum и praevomer. По ходу бороздки и распределению лучей шагрени мы можем определить, что центр окостенения praevomer находился у самого переднего края кости, что имеет значение для восстановления характера черепа в более молодом возрасте или у предковой формы.
Зубы челюстного ряда по типу своего прирастания имеют отчасти плевродонтный характер, однако, в общем мало отличаются от зубов многих стегоцефалов. Исследование поперечных разрезов одного зуба, выполненное но моей просьбе А. П. Быстровым, показало, что они отличаются от зубов стегоцефалов полным отсутствием складок на дентинной стенке. Зубы построены как зубы рептилий.

Рис. 3. Lanthanosuchus watsoni, gen. et sp. nov. A — часть зубного ряда верхней челюсти, увеличено; B — quadratojugale dexirum, снизу; C — очертания septomaxillare, сверху; D —центральная часть затылка (с черепа № 271/1). Обозначения как на рис. 1 и 2 и еще: X — XII — отверстие блуждающего и подъязычного нервов; F. pt — посттемпоральное окно.
Затылочная поверхность черепа (рис. 3, D и 4) характерна сильной уплощенностью, а также выдвинутыми назад костями крыши черепа, затылочный край которых отвернут кверху и осложнен разнообразными костными выростами. Центральная часть затылка выделяется своей компактностью, а также тем, что она имеет наклонную поверхность, покатую назад. Поэтому центральная часть затылка выдвинута из-под навеса крыши черепа в каудальном направлении. Эта часть затылка, ввиду важности своих деталей, изображена отдельно (рис. 3, D).
Basioccipitale, составляющее основу центральной части затылка, уплощенное, поперечно-овальное, имеет глубокую ямку для хорды. Боковые оконечности basioccipitale выступают наружу и снизу прикрываются широкими лепестками задних углов парасфеноида, простирающимися латерально в виде свободно торчащих пластинок.
Exoccipitalia опираются на basioccipitale своими нижними колоннообразными частями и почти соприкасаются по средней линии, отделяя foramen magnum от basioccipitale. Выше exoccipitalia сильно расходятся в стороны, занимая косое положение, их верхние части расширены и сочленяются своими внутренними углами с supraoccipitale и с paroccipitalia по верхнему краю. В середине последнего шва находятся углубление и широкий просвет fenestra posttemporalis.
Supraoccipitale — выпуклая пластинка субквадратной формы, расположенная наклонно и покрывающая собою значительный участок заднего отдела мозговой коробки. Нижний край кости ограничивающий foramen magnum, утолщен и от него до середины кости проходит заметный гребень. Боковые края supraoccipitale сочленяются с exoccipitalia внизу и с paroccipitalia снаружи. Передне-верхний край supraoccipitale целиком соединен с parietalia.
Рагoccipitalia — массивные кости, составляющие непосредственное продолжение вверх расходящихся exoccipitalia образуют наружно-верхние углы центральной части затылка и сочленяются с parietalia изнутри, с dermosupraoccipitalia вверху и с tabularia на супралатеральных оконечностях.
Exoccipitalia прилегают сзади к paroccipitalia, которые впереди первых образуют часть задней стенки мозговой коробки. Нижние части paroccipitalia впереди exoccipitalia спускаются до basioccipitale.
Посттемпоральные окна (fenestra posttemporalis) для латеральных головных вен (v. capitis lateralis) очень узки и проходят под paroccipitalia, между последними и exoccipitalia, совсем иначе, чем у большинства стегоцефалов, и совершенно так же, как у Anthracosaurus и Seymouria. Отверстие X нерва большое и находится между передней стороной базальной части каждого exoccipitale и нижней частью рагосcipitale, даже частично прилегая к наружной стенке basioccipitale. В медиальной стенке задней части отверстия X нерва имеется маленькое отверстие XII нерва (for. n. hypoglossi).
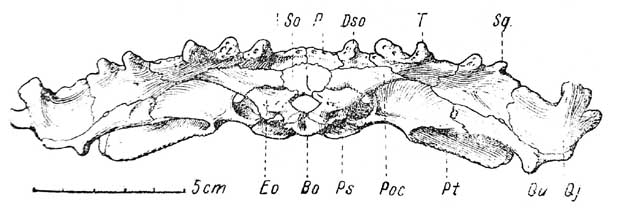
Рис. 4. Lanthanosuchus watsoni. gen et sp. nov. Череп № 271/1, затылочная поверхность. Обозначения как на предыдущих рисунках.
Pterygoidea на затылочной поверхности показывают высокие восходящие пластинки (laminae ascendentes), достигающие крыши черепа и несущие на задних сторонах воронкообразные углубления, подобно области среднего уха у стегоцефалов.
Однако птеригоиды в затылочном отделе отодвинуты далеко наружу от центральной части затылка с мозговой коробкой. Для суждения о строении среднего уха у нашей формы не имеется достаточных данных, так как от обеих stapes не осталось никаких следов и на неполно окостеневшей мозговой коробке нельзя установить точное местоположение fenestra ovalis.
Общее строение затылочного отдела дает все же основания предположить, что среднее ухо было построено скорее по типу рептилий, чем стегоцефалов, и stapes должен был занимать близкое к горизонтальному положение. Это предположение подкрепляется наличием ямки на задне-внутренней стороне медиального отростка quadratum, в которую мог упираться дистальный конец stapes.

Рис. 5. Lanthanosuchus watsoni. gen et sp. nov. Череп № 271/1.
A — вид сбоку; B — вид сбоку при мысленно разрезанном парасагитально черепе. Обозначения как на предыдущих рисунках и еще: Ep —epipterygoideum; l. a. — восходящая пластинка птеригоида (lamina ascendens). Орбитосфеноид не показан.
Мозговая коробка (рис. 5, В) слабо окостеневала. Костное дно мозговой коробки образовано парасфеноидом, так как basisphenoideum полностью оставался хрящевым. Из латеральных элементов окостененел только эпиптеригоид, построенный очень просто, в виде плоской костной палочки, слегка расширяющейся вверху и внизу. Орбитосфеноид короткий, окостеневал сравнительно слабо (на рисунке не показан, для наглядности характеристики внутренней полости черепа).
Череп необыкновенно сильно уплощен в преорбитальном отделе (рис. 5, А и В).
В области хоан кости небной поверхности непосредственно соприкасаются с костями крыши черепа. При жизни животного между ними должен был существовать какой-то просвет, но этот просвет, во всяком случае, был поразительно малым.
Назад внутренняя полость черепа постепенно повышается и достигает своего максимума в париетальном отделе. Однако довольно большие глаза животного должны были очень сильно выступать наружу, несмотря на резко поднятые края орбит.
Найденные отдельно при раскопках 1934—1935 гг. позвонки, а также три неполных позвонка, найденные вместе с черепом в 1939 г., чрезвычайно сходны с позвонками Seymouria. Сходство настолько велико, что эти позвонки могли бы быть отнесены к последней. В общем позвонки нашей формы крупнее и несколько массивнее позвонков сеймурии, обычного для рептилий гастроцентрального типа, со следами больших интерцентров. Позвонки обладают очень широкими и массивными невральными дугами и зигапофизами. Остистые отростки сравнительно тонкие и не имеют на верхних концах массивных утолщений (Ефремов, 1940, табл. VI, фиг. 1, 2, 3).
Для характеристики позвонков сеймуридного типа могут служить прекрасные изображения позвонков Kotlassia, данные А. П. Быстровым (Bystrow, 1944), одно из которых здесь приведено (рис. 6).

Рис. 6. Kotlassia prima Amal. Позвонки. [По Быстрову (Bystrow), 1944].
Цифры, стоящие над позвонками, соответствуют порядковому номеру каждого изображенного позвонка.
Изложенное в общем исчерпывает материалы, имеющиеся по вновь открытой форме. Мы не можем составить себе представления о посткраниальном скелете животного, хотя общность строения его позвоночного столба с такой формой, как Seymouria, у которой посткраниальный скелет хорошо известен, позволяет предположить, что скелет нашей формы в общих чертах мог быть подобен скелету Seymouria.
Строение новой формы настолько необычайно, что без всякого сомнения наша форма представляет собою новый род и вид. В качестве родового названия необходимо оставить уже опубликованное мною (Ефремов, 1941) название Lanthanosuchus.
Видовое название L. qualeni установлено мною по фрагментам крыши черепа (№ 157/209), найденным в раскопках 1934—1935 гг. Сравнение этих фрагментов с описываемым черепом показало некоторые отличия в размерах, форме, характере гребней и скульптуры, могущие быть видовыми отличиями. Поэтому вновь найденный череп, вероятно, принадлежит к другому виду и должен получить новое видовое название, которое я предлагаю в честь крупнейшего палеонтолога — исследователя древнейших Tetrapoda профессора D. M. S. Watson.
Таким образом полное название новой формы — Lanthanosuchus watsoni, gen. et sp. nov. Вопрос о принадлежности Lanthanosuchus к более высоким систематическим категориям не может быть решен так просто, хотя необычайность его строения вполне обосновывает выделение нового семейства, подотряда и даже отряда. Рассмотрим вкратце морфологические особенности Lanthanosuchus.
Животное обладает архаическим общим строением черепа. Широкая параболическая форма черепа свойственна наиболее примитивным стегоцефалам и, главным образом, личинкам последних. Строение небной поверхности черепа также крайне примитивно и характерно для древнейших эмболомерных стегоцефалов, равно как и наличие суборбитальных ям.
Отсутствие дополнительных зубных рядов и околохоанных больших зубов не установлено ни у одного из известных стегоцефалов, в то время как общий вид неба с его радиальной шагренью имеет прямо-таки рыбий характер.
Вместе с тем ряд признаков лантанозуха свойствен уже только поздним рахитомным стегоцефалам — слабое окостенение мозговой коробки, развитие corpus parasphenoidei, сочленение птеригоидов непосредственно с парасфеноидом, крайнее уплощение черепа в степени, наблюдающейся только у последних триасовых Stereospondyli.
В этот смешанный комплекс стегоцефальных признаков вплетены еще более многочисленные признаки примитивных рептилий. Строение ноздрей и хоан, структура зубов и костей крыши черепа, положение отверстий X и XII нервов, ограничение орбит, большие размеры париетального отверстия, характер слухового отдела черепа, наличие orbitosphenoideum, строение и соотношения supraoccipitale, развитие quadratum, палочковидный epipterygoideum, не говоря уже о типично рептильных позвонках котилозаврового типа, — все эти признаки характерны для рептилий, но отнюдь не для амфибий.
Особый характер имеет необыкновенная скульптура и развитие костных выростов на крыше черепа — это признаки особенной адаптации Lanthanosuchus, равно как и сильное развитие quadratojugalia. Характер скульптуры объясняется необходимостью повышения механической прочности крайне уплощенного черепа, примитивное небо которого переложило всю нагрузку на крышу черепа. Остальные особенности из указанных выше пока еще не могут быть с достоверностью объяснены.
Крайне характерно расположение посттемпоральных окон в затылке лантанозуха. Эти отверстия, очень узкие, проходят между paroccipitalia (opisthotica) и exoccipitalia, что не наблюдается у стегоцефалов (за исключением карбонового Anthracosaurus) и очень типично для Seymouria. Такое же сходство Lanthanosuchus с Seymouria и Anthracosaurus наблюдается в месте выхода X нерва. Foramen nervi vagi располагается между exoccipitale и paroccipitale, а не пронизывает exoccipitale, как у всех стегоцефалов.
Расположение каналов внутренних сонных артерий у лантанозуха крайне своеобразно. Они не заключены в кости парасфеноида, как у стегоцефалов, и подходят непосредственно к basisphenoideum, как у рептилий. Однако они лежат впереди сочленения птеригоидов с basis cranii, как это наблюдается у стегоцефалов. В то же время это сочленение отнесено у лантанозуха далеко назад, и имеется второе соединение птеригоидов с передней частью corpus parasphenoidei, относительно которого каротидные отверстия оказываются расположенными сзади. Таким образом характер каналов art. carotidis internae у Lanthanosuchus можно рассматривать и как амфибиальный, и как рептильный.
Наконец, совсем особенное значение в анализе строения лантанозуха имеет наличие больших височных ям, сопровождающихся развитием птеригоидных флангов, так же как это наблюдается у тероморфных рептилий.
Находка Lanthanosuchus лишний раз свидетельствует, что развитие и положение височных ям, считавшееся одним из важнейших таксономическнх признаков классификации отрядов рептилий, на самом деле является сравнительно пластическим признаком, связанным с определенной функцией челюстей и расположением их аддукторных мышц.
Развитие височных ям лантанозуха является несомненно прогрессивным признаком, появившимся вместе с тем на чрезвычайно архаической основе. Височные ямы лантанозуха указывают на сравнительно большую массу аддукторных мышц нижней челюсти. О том же говорят сильные поперечные фланги птеригоидов, служащие для поддержки мышц птеригоидной группы. В этом отношении строение височной области Lanthanosuchus аналогично таковому терапсидных рептилий. Вероятно, нижняя челюсть лантанозуха должна значительно отличаться от примитивных стегоцефалообразных челюстей древнейших рептилий и, возможно, обладала развитым венечным отростком.
Однако птеригоидные фланги лантанозуха отодвинуты далеко назад по сравнению с Therapsida и находятся под центром височной ямы, а не у ее переднего края, как у последних. Поэтому общая схема тяги челюстных мышц у лантанозуха была иной, чем у рептилий. Несомненно, Lanthanosuchus обладал гораздо более слабыми темпоральными мускулами, и вертикальная тяга челюсти осуществлялась, главным образом, за счет подвинутых назад птеригоидных мышц, прикреплявшихся к нижнемедиальному краю нижней челюсти и подменявших группу мышц, опирающуюся на мозговую коробку у прогрессивных рептилий.
Поэтому, поскольку у Lanthanosuchus отсутствует важнейший прогрессивный признак эволюции синапсидных рептилий — перенесение опоры аддукторных мышц челюсти на мозговую коробку, не исключена возможность, что прободение мышцами крыши черепа и образование височной ямы отчасти обязаны уплощению черепа в процессе эволюции лантанозухов, а не только увеличению мышечной массы и размаха конечных отростков, как это имеет место у зверообразных рептилий.
Причисление Lanthanosuchus к синапсидным рептилиям в качестве особой уклонившейся формы явилось бы ошибкой. Височные ямы нашей формы могут рассматриваться только как своеобразное проявление прогрессивного признака на весьма архаической основе, качественно отличного от настоящих височных ям Synapsida. В самом деле, височная яма Lanthanosuchus, не имеющая в качестве главной опоры стенок мозговой коробки, с преобладающей ролью птеригоидных мышц, сходна с височной ямой синапсид преимущественно внешне и отличается, так сказать, другим содержанием.
Подробный анализ анатомического строения Lanthanosuchus подводит нас к выводам иного характера.
Лантанозух является по совокупности своих признаков рептилией, оправленной в архаическую форму стегоцефального черепа, удержавшей ряд весьма древних черт строения эмболомерных амфибий карбона, вместе с тем приобретшей уплощение черепа и упрощенную мозговую коробку поздних стегоцефалов, наряду с развитием височных ям.
Таким образом в Lanthanosuchus мы видим смешение признаков амфибий и рептилий в их наиболее примитивном выражении и, с другой стороны, смешение путей развития, свойственных гораздо более поздним группам рептилий и стегоцефалов, окончательно определившихся в совершенно различных направлениях эволюции. Нужно думать, что лантанозух является представителем какой-то древней группы Tetrapoda, реликтом того времени, когда совершалось превращение еще тесно связанных с водой древних стегоцефалов во вполне наземных рептилий и когда возможные направления эволюции еще не были сведены естественным отбором в основные стволы развития рептилий, все дальше уклонявшиеся от своих предков — стегоцефальных амфибий. Нельзя забывать, что лантанозух найден в нижних горизонтах верхней перми; самое существование столь архаической формы среди прогрессивных гондванских зверообразных рептилий Ишеевской фауны можно объяснить только реликтовым ее характером, обитанием в особой экологической нише, исключавшей возможность постоянного соприкосновения с новыми элементами фауны. В самом деле, уже один только внешний облик лантонозуха говорит о том, что его экологический тип должен был быть своеобразным. Животное имело странный вид со своими костными выростами и большими глазами, уродливо выпяченными из необычайно плоского черепа.
Для решения вопроса о том, к какой же группе может принадлежать Lanthanosuchus, следует обратиться к другим формам, обладающим переходным между Amphibia и Reptilia характером строения.
В 1896 г. знаменитый американский палеонтолог Коп (Соре, 1896) описал из нижнепермских красноцветов Тексаса неполно сохранившийся череп небольшого животного, названного им Conodectes. Коп отнес новую форму к группе котилозавров — пресмыкающихся со сплошной крышей черепа, и считал ее за вероятного предка черепах. Неполная сохранность и обычная для ранних палеонтологических работ недостаточная детальность описания, оставили незамеченными некоторые интересные особенности Conodectes.
Из тех же отложений Броили (Broili, 1904) описал небольшое пресмыкающееся, очень близкое к Conodectes, получившее название сеймурии (Seymouria). Сеймурия возбудила среди палеонтологов живейший интерес. В строении скелета сеймурии оказались настолько смешанными анатомические признаки амфибий и рептилий, что в течение всех последующих лет видные палеонтологи мира пытались решить вопрос о том, куда же, к какому классу следует отвести сеймурию — к амфибиям или рептилиям. Нужно оговориться, что здесь речь идет не о современных, далеко дивергировавших представителях обоих классов, а о древнейших, наиболее примитивных группах земноводных и пресмыкающихся, обладающих гораздо более сходным, чем современные, строением.
Уотсон (Watson, 1918) в нескольких работах, изучив подробно скелет сеймурии, высказался за принадлежность ее к котилозаврам, согласно с Броили. Брум (Broom, 1913) отнес сеймурию к эмболомерным стегоцефалам, а Сушкин (Sushkin, 1925, 1927) на основании анализа ее мозговой коробки и, особенно, слухового отдела черепа показал, что по этим признакам сеймурия есть несомненная амфибия.
Наконец, в 1939 г. появилась прекрасная работа Уайта (White, 1939) с весьма детальным исследованием скелета Seymouria по массовому материалу американских музеев. Уайт решил, что, хотя сеймурия обладает многими анатомическими признаками амфибий, все же ее причисление к рептилиям имеет более веские основания. Рассмотрим вкратце важнейшие особенности этого загадочного животного.

Рис. 7. Seymouria baylorensis Broili. Реконструкция крыши черепа. [По Уайту (White), 1939]. 2/3 натуральной величины.
Обозначения в общем соответствуют данным на рис. 1—4, только отдельные индексы имеют дальнейшие буквы названия кости. Кроме того: Smx — septomaxillare; It — intertemporale. Пунктиром обозначен ход ductus nasolacrymalis.
Сеймурия — небольшое животное длиной около одного метра от морды до конца хвоста. Череп ее (рис. 7) со сплошной крышей покровных костей характерен наличием intertemporale. Как в крыше, так и в эндокраниальном отделе имеется весь комплекс костей, характеризующий черепа наиболее древних эмболомерных амфибий нижнего карбона. Глубокие и длинные слуховые вырезки построены по стегоцефальному типу, отдел среднего уха представляет собою почти вертикальную воронку с направленными косо вверх к центру слуховой вырезки stapes, что является характерным признаком всех стегоцефальных амфибий. Ноздри сближены и внутри их сильно развиты septomaxillaria, что очень сходно с примитивными рептилиями и, в частности, с высшими котилозаврами. Кости черепа покрыты сетчатой нерезкой скульптурой, каналы боковой линии совершенно отсутствуют. Подобная скульптура, кроме стегоцефалов, известна у некоторых настоящих котилозавров — Сарthorhinus и Labidosaurus. Небная поверхность черепа (рис. 8) сплошь закрыта костями, как у эмболомерных стегоцефалов, однако хоаны сближены между собою так, как это наблюдается у примитивных рептилий. Наружные края птеригоидов отвернуты вниз и отделены от стенок черепа; они образуют птеригоидные фланги, очень характерные для всех древних рептилий. Парасфеноид велик и широк, с коротким, как у Embolomeri, передним отростком и закрывает снизу дно мозговой капсулы, которое у сеймурии не окостеневает, как у поздних стегоцефалов, и, напротив, полностью окостеневает у древнейших амфибий и у котилозавров (так же, как у всех рептилий). Каналы внутренних сонных артерий пронизывают окостеневшую часть базисфеноида непосредственно сзади базиптеригоидных отростков, что характерно для рептилий. Вместо сплошного окостенения sphenethmoideum, типичного для стегоцефалов, сеймурия имеет парные орбитосфеноиды, подобно примитивным рептилиям. Эпиптеригоид широкий и пластинчатый, амфибиального характера. В паротикальном отделе черепа Seymouria мы встречаем вертикальный гребень, кольцевой канал вокруг проксимальной части stapes и другие признаки, известные у кроссоптеригий (Megalichthys) и утерянные у большинства древнейших амфибий.

Рис. 8. Seymouria baylorensis Broili. Реконструкция вентральной поверхности черепа [По Уайту (White), 1939]. 2/3 натуральной величины.
Обозначения как на прежних рисунках, за исключением: Bpt — базиптеригоидные отростки; Ecpt — transversum (ectopterygoideum); Opis — paroccipitate (opisthoticum).
Очень интересной деталью в затылочном отделе черепа сеймурии является сплошное соединение paroccipitale с tabulare, вследствие чего посттемпоральные окна для латеральных головных вен проходят под paroccipitalia, а не над ними, как у всех стегоцефалов. Такое строение, как верно отмечено Уайтом, очень характерно для Anthracosaurus — особенного эмболомера из нижнего карбона Шотландии, считающегося представителем наиболее близкой рептилиям ветви эмболомерных стегоцефалов.
Отверстие X нерва у сеймурии проходит не сквозь exoccipitale, а между этой костью и paroccipitale, что также характерно для Anthracosaurus. Нижняя челюсть Seymouria имеет полностью стегоцефальный тип строения; зубы со слабо складчатым дентином, как у древнейших Embolomeri.
Скелет построен целиком по типу примитивных котилозавров. Позвонки обычного для рептилий гастроцентрального типа сохраняют огромные интерцентры и обладают очень широкими и массивными невральными дугами, столь характерными для всех котилозавров. Атлас и аксис неполно окостеневают снизу, что совершенно необычно для рептилий.
Двухголовчатыс передние ребра дистально расширены.
Уайт подчеркивает, что Seymouria имеет много черт специализации, по которым эта форма не может быть предком ни одного из известных в настоящее время котилозавров. Так, например, ширина ее невральных дуг значительно превосходит (в полтора раза) наиболее широкие невральные дуги котилозавров. Отмечено усиление этого признака с возрастом. Широкие невральные дуги развиваются для фиксации средней части позвоночного столба, для сохранения равновесия тела во время ходьбы при столь характерном для котилозавров слабом развитии илио-феморальных (ягодичных) мускулов, т. е. именно тех мышц, которые помогают сохранять равновесие при ходьбе.
Уайт делит котилозавров по этому признаку на две главные линии эволюции — бòльшая часть обладает широкими невральными дугами и зигапофизами и слабыми илио-феморальными мускулами, другие (Solenodosaurus из верхнего карбона Чехословакии) имеют, подобно высшим группам рептилий, сильные мышцы и более узкие невральные дуги. С точки зрения этой адаптации сеймурия представляет собою крайний пример специализации первой линии котилозавров.
Спор о принадлежности Seymouria к амфибиям или рептилиям в конце концов не мог бы быть разрешенным с достаточной достоверностью, если бы сеймурия представляла собою единичное уклонение архаического типа. Но на территории нашего Союза были найдены остатки подобных же животных, также обладающие смешанными признаками амфибий и рептилий. В начале нашего века В. П. Амалицкий открыл в верхнепермских отложениях Сев. Двины среди массы остатков гондванских зверообразных рептилий два неполных скелета небольших стегоцефалообразных животных, близких к Seymouria и Conodectes. Краткие диагнозы новой формы, получившей название котлассии (Kotlassia), были опубликованы после смерти Амалицкого в 1922 г. Амалицкий считая котлассию подродом сеймурии — так поразило его внешнее сходство обеих форм. Сушкин в нескольких работах 1925, 1927 и 1928 гг. дал подробное описание котлассии и выяснил существенные отличия ее от сеймурии, поместив обе формы в группу сеймуриаморф, принадлежащую к амфибиям. Окончательно освещение котлассия получила только после детальной препаровки и превосходного описания А. П. Быстрова, напечатанного, с задержкой из-за войны, только в 1944 г. (Bystrow, 1944).
Общий характер скелета котлассии тот же, что у сеймурии. Позвонки обеих форм почти неотличимы, за исключением более высоких и массивных у котлассии остистых отростков и сращения интерцентров с центрами в поясничных позвонках котлассии (рис. 6). Кости конечностей котлассии более удлиненные и не так массивны, как у сеймурии. Котлассия обладает костным спинным щитом, составленным из отдельных сильно орнаментированных элементов, оригинальной балочной скульптуры. В связи с этим концы остистых отростков позвонков расширены и утолщены.
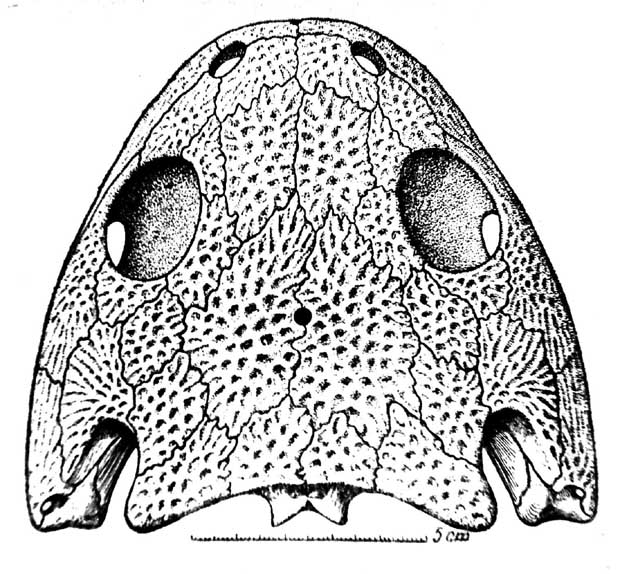
Рис. 9. Kotlassia prima Amal. Реконструкция крыши черепа. [По Быстрову (Bystrow), 1944].

Рис. 10. Kotlassia prima Amal. Реконструкция вентральной поверхности черепа. [По Быстрову (Bystrow), 1944).
Обозначения как прежде, за исключением: Fo — fenestra ovalis; St — stapes; V — praevomer.
В черепе котлассии различия обеих форм проявляются значительно резче. Параболический череп котлассии несет более ярко выраженный стегоцефальный характер (рис. 9). Слуховые вырезки мельче и сходны с таковыми у поздних рахитомных стегоцефалов. Ноздри округлые и широко раздвинуты, без заметного septomaxillare. На краю морды характерное для стегоцефалов foramen intermaxillare. Хоаны котлассии (рис. 10) разделены широкими praevomera, как у всех стегоцефалов. Позади них имеется дополнительный зубной ряд, проходящий по небной и поперечной костям, в то время как у сеймурии, в отличие от стегоцефалов, имеются только по четыре зуба в окружении каждой хоаны. Передний отросток парасфеноида гораздо длиннее, чем у сеймурии, и вклинивается между превомерами, как у некоторых рахитомных стегоцефалов. Имеются зачаточные межптеригоидные полости, поперечные фланги птеригоидов хотя и присутствуют, но развиты значительно слабее, чем у сеймурии. Sphenethmoideum окостеневает сплошной костью, как у стегоцефалов. Посттемпоральные окна проходят под paroccipitalia, как у всех стегоцефалов, но отверстие X нерва остается в том же положении, как у сеймурии (рис. 11). Передняя часть базисфеноида (sella turcica) — окостеневает сильнее, чем у сеймурии. В области челюстного мыщелка у Kotlassia имеется for. рагаquadratum, характерное для стегоцефалов. Это показывает, что устройство челюстей у Kotlassia было вполне амфибиальное, чем она отличается от сеймурии и, конечно, от лантанозуха. В остальном строение черепа котлассии повторяет таковое сеймурии. Нижние челюсти обеих форм крайне сходны. Зубы котлассии имеют тот же складчатый лабиринтодонтный характер.

Рис. 11. Kotlassia prima Amal. Реконструкция затылочной поверхности черепа. [По Быстрову (Bystrow), 1944]. 2/3 натуральной величины.
Обозначения как прежде и еще: F. pq. — foramen paraquadratum; IX—X — отверстие языкоглоточного и блуждающего нервов.
Таким образом котлассию нужно рассматривать как подобную сеймурии форму, но обладающую несколько более амфибиальным характером строения, более амфибию, так сказать.
Когда я консультировал работу А. П. Быстрова, он долго был в нерешительности, куда отнести котлассию по данным анатомического строения скелета, поскольку в его руках не было решающих данных — проходила котлассия подобно всем амфибиям метаморфоз или размножалась, как рептилия. За отсутствием возможности кардинального решения пришлось разбить основные анатомические признаки котлассии, которых можно было насчитать 46, на две группы — признаки амфибиальные и признаки рептильные. Первых оказалось 24, вторых 22, и Быстров отнес котлассию к амфибиям, тем более, что в числе первых признаков были такие важные, как строение слухового отдела.
Таким образом из двух весьма близких форм в результате детальнейшего изучения одна была причислена к рептилиям, другая — к амфибиям. Это обстоятельство говорит о наличии какой-то промежуточной группы, переходной между двумя классами, отдельные представители которой могут иметь преобладание тех или иных признаков, как бы сгущение их в сторону Amphibia или в сторону Reptilia. Для создания более полного представления об этой группе решающим оказалось открытие Lanthanosuchus. Лантанозух обладает сходством с Seymouria и Anthracosaurus по своеобразному строению затылочного отдела черепа, что может служить указанием на общность их происхождения. Лантанозух имеет более ярко выраженные признаки рептильного характера в строении затылка, слухового отдела и в наличии височных ям, чем сеймурия и, тем более, котлассия. Вместе с тем в окостенении мозговой коробки и в развитии парасфеноида он ближе к амфибиям, чем котлассия, не говоря уже о сеймурии. Зубы Lanthanosuchus уже утеряли складчатость дентина — наследие стегоцефалов в зубах сеймурии и котлассии, однако строение его небной поверхности примитивнее, чем у двух этих форм. И сеймурия, и лантанозух утеряли foramen interpraemaxillaris, и, следовательно, якобсонов орган, котлассия же сохранила его. Наряду с этим, при стегоцефальном характере затылка, череп сеймурии весьма сходен с черепом котилозавров, а лантанозух приобрел такое уплощение черепа, которое свойственно только последним стадиям эволюции стегоцефалов. Крайнее сходство позвонков Lanthanosuchus, Seymouria и Kotlassia, разумеется не случайно и указывает, с одной стороны, на родство всех трех форм, с другой — на общий тип строения скелета и, следовательно, на общий тип движения.
Скелет лантанозуха нам пока неизвестен. Скелет котлассии отличается очень массивным плечевым поясом, придвинутым близко к черепу, как это наблюдается у всех стегоцефалов. У сеймурии, хотя реконструкция Уайта не кажется мне правильной (плечевой пояс сдвинут слишком далеко назад и его вентральная часть слишком выдвинута вперед, вследствие чего шейный отдел Seymouria сделан подобным тероморфной рептилии, intеrclavicula наклонена и удалена от ребер, конечности слишком высоко подняты) и придает ей преувеличенное сходство с рептилиями, плечевой пояс значительно более облегчен и подвижен. Ольсон (Olson, 1936), анализировав аксиальную мускулатуру котилозавра Diadectes пришел к заключению, что, несмотря на тяжелый плечевой пояс, Diadectes и Seymouria были способны к реберному дыханию, составлявшему одно из главных приобретений и преимуществ класса рептилий. По всей вероятности, у сеймурии, как и у наиболее примитивных котилозавров, существовали одновременно оба способа дыхания — дном ротовой полости, как у амфибий, и реберное дыхание, причем первый способ несомненно преобладал. Котлассия, с более массивным стегоцефалообразным плечевым поясом, придвинутым к черепу, могла иметь реберное дыхание лишь в зачатке и дышала в общем подобно стегоцефалам. Повидимому, в прямой связи с этим стоит сохранение котлассией якобсонова органа (широко раздвинутые ноздри и хоаны, наличие for. interpraemaxillare). Если это сопоставление правильно, то лантанозух со своими ноздрями рептильного типа должен был иметь, подобно сеймурии смешанное дыхание.
Вопрос о характере размножения, остающийся совершенно открытым для лантанозуха, для сеймурии и котлассии также далек от разрешения. Некоторые косвенные указания дает Уайт при анализе полового диморфизма в скелете Seymouria. Уайт находит эти половые различия похожими на имеющиеся в скелетах современных крокодилов и черепах (перемещение шевронов и т. д.). Это дает возможность предположить наличие внутреннего оплодотворения у сеймурии и, следовательно, наличие богатых желтком яиц и безметаморфозного размножения. Котлассия, по большому сходству строения своего скелета с сеймурией, может также обладать сходным способом размножения. Правда, скелет котлассии в общем менее массивен и крепок, чем скелет сеймурии, и это служит указанием на более близко связанный с водой образ ее жизни.
Итак, мы рассмотрели три формы, строение которых известно теперь с достаточной полнотой. И сеймурия, и лантанозух, и котлассия — все несомненно родственны между собою. Признаки, их объединяющие, можно разделить на две категории.
В первую войдут весьма архаические черты, свойственные древнейшим эмболомерным амфибиям. В другую должны быть отнесены признаки, характерные для наиболее примитивных рептилий, древнейших котилозавров, как, например, общий для всех трех форм тип строения позвоночника, безусловно не амфибиального.
Обе эти категории признаков смешаны в строении всех трех форм приблизительно в равных пропорциях и определяют промежуточный между двумя классами характер рассматриваемой группы.
Сеймурия, лантанозух и котлассия по этим двум категориям признаков представляют собою, грубо говоря, смесь эмболомерного стегоцефала и архаического котилозавра. Но каждая из форм имеет еще свои собственные черты специализации в том или другом направлении, накладывающиеся на общий тип строения и осложняющие анализ его при изучении каждой формы в отдельности.
Сравнение сеймурии и лантанозуха, например, показывает далеко зашедшую дивергенцию, выраженную в весьма крупных изменениях структуры черепа. Все это убедительно говорит о том, что сеймурия, лантанозух и котлассия принадлежат к какой-то очень древней группе форм, обладавших промежуточным между древнейшими амфибиями и рептилиями строением, группе, вероятно, богатой видами, отдельные ветви которой значительно дивергировали и уклонялись.
В самом деле, уже только в трех изученных нами формах мы видим разные приспособления, касающиеся весьма важных биологических систем органов (дыхание сеймурии, височные ямы лантанозуха), появившиеся вместе с такими адаптивными признаками, которые характерны для форм, прошедших совершенно иные пути эволюционного развития (уплощение черепа, редукция костной мозговой коробки лантанозуха). Котлассия, например, сохраняя амфибиальный тип дыхания, более близка к сеймурии и к котилозаврам по мозговой коробке, чем опередивший ее по ряду признаков рептилий лантанозух. Все подобные явления легко объяснимы предположением о существовании в ранние периоды эволюции древнейших Tetrapoda большой группы форм, адаптивная радиация которой, так оказать, «нащупывала» различные пути эволюции. В этой группе на весьма архаической основе возникали различные специализации, в известном смысле предварявшие прогрессивные достижения позднейших эволюционных стволов, но быстро уничтожавшиеся естественным отбором, так как общий уровень организации животных не мог доставить им серьезных преимуществ в сравнении со стегоцефалами.
С точки зрения наличия подобной группы понятно и строение лантанозуха — «почти рептилии», эволюционировавшей как обыкновенный стегоцефал, и сеймурии — «почти стегоцефала», эволюционировавшей как рептилия, и котлассии, занявшей промежуточное между ними положение в смысле своей биологии. Развитие этой промежуточной группы должно было начаться раньше появления настоящих рептилий, т. е. приблизительно в середине карбона.
Повидимому, наиболее древним известным нам членом этой группы является эмболомерный стегоцефал нижнего (вернее низов верхнего карбона, при двучленном делении) карбона, Anthracosaurus, не случайно постоянно выделявшийся исследователями в особую эволюционную линию древних амфибий, получившую у Севе-Зедерберга название Reptiliomorpha. Я указывал уже на несомненное родство Antnracosaurus и наших трех форм.
Все перечисленные члены промежуточной группы разновозрастны. Антракозавр найден в нижнем карбоне Англии, сеймурия с близким ей конодектесом в красноцветах низов перми Сев. Америки, лантанозух в нижних горизонтах верхней перми Советского Союза, а котлассия в ее верхних горизонтах.
Все рассмотренные формы являются редкими, единичными находками среди огромного количества остатков более прогрессивных амфибий и рептилий при крупных раскопках. Каждая из этих форм повидимому, занимает особую экологическую нишу, какую именно еще трудно себе представить, но, во всяком случае, резко отличающуюся от всех других, найденных совместно форм той или другой фауны.
Сеймурия среди водных рыбоядных пеликозавров, роющих котилозавров и крупных стегоцефалов фауны красноцветов Тексаса, повидимому, обитала на суше, укрываясь среди болотной растительности. Сеймурия питалась, повидимому, мелкой пищей из наземных и речных моллюсков, червей, насекомых, мальков рыб, стегоцефалов и современных ей рептилий. Котлассия с ее более амфибиальным обликом в биоценозе гондванских поздних рептилий, видимо, аналогична панцырным роющим стегоцефалам красноцветов Тексаса — так называемым диссорофидам. Питание котлассии было в общем сходно с питанием сеймурии, но с несомненным преобладанием рыбной пищи, судя по озубленню. Лантанозух, своим уплощенным черепом предвосхитивший гораздо более позднюю специализацию триасовых стегоцефалов — брахиопид, выполнял роль малоподвижного живого капкана, охотясь за мелкой рыбой в небольших рукавах речных дельт, куда не заходили огромные титанозухиды из рептилий или длинномордые, гавиалообразные стегоцефалы-платиопсы, составлявшие единый фаунистический комплекс.
Лантанозух питался преимущественно рыбой, вернувшись назад в этом отношении от сеймурии, чем и объясняется стегоцефальный тип его адаптации.
По всем данным, сеймурия, лантанозух и котлассия представляют собою редкие реликты, пережитки древней группы, некогда гораздо более многочисленной. Благодаря своей оригинальной приспособленности эти реликты могли войти в состав биоценозов мощной пермской адаптивной радиации рептилий и амфибий и сохраниться в них до очень позднего времени со всеми своими архаическими чертами строения, поражающими исследователя на фоне уже далеко продвинувшейся общей эволюции сопутствующих форм.
Как только мы делаем попытку выяснить систематическую принадлежность сеймурии, лантанозуха или котлассии к тому или другому классу, не вырывая отдельных, может быть, зачастую не вполне веских признаков, так сразу же теряем под ногами почву. В этом нет ничего удивительного, так как вся группа в целом, остатками которой эти три формы являются, есть группа переходная, промежуточная.
Поскольку в этой группе мы видим смешение важнейших признаков обоих классов и отдельные, встречающиеся в ней обособления заслуживают выделения крупных систематических категорий типа, отрядов, нужно думать, что группа в целом по своему значению в эволюции древнейших четвероногих имеет характер подкласса.
Я предлагаю для этого подкласса название Batrachosauria — лягушкоящеров, отражающее промежуточное, переходное между обоими классами значение батрахозавров, хотя с таким же правом мы можем назвать его Saurobatrachia, имея в виду, например, лантанозуха.
Вопрос о причислении батрахозавров к тому или другому классу не имеет существенного значения и может быть решен в ту или другую сторону только в случае получения достоверных данных о характере размножения или кожных покровов представителей нового подкласса.
Не подлежит никакому сомнению, что при дальнейших исследованиях число членов подкласса батрахозавров будет увеличиваться. Весьма вероятно, что к батрахозаврам принадлежат еще недостаточно изученные формы Cardiocephalus из красноцветов Тексаса, Eosauravus из верхнего карбона Виргинии, Solenodosaurus из верхнего карбона Чехословакии. Мелкие среднекарбоновые микрозавры типа тудитанид, обладающие плечевым поясом рептилийного типа, при получении экземпляров лучшей сохранности, вероятно, также войдут в состав подкласса батрахозавров. В пределах Советского Союза имеются определенные указания на наличие новых форм батрахозавров в самых низких горизонтах верхней перми — I зоне.
В свете сказанного, систематическое положение нашей новой формы Lanthanosuchus watsoni может быть представлено следующим образом: подкласс Batrachosauria, отряд (условно) Seymouriamorpha, сем. Lanthanosuchidae nov., род Lanthanosuchus. Выделение отрядов в новой группе батрахозавров представляется мне преждевременным и может быть сделано лишь впоследствии, когда в нашем распоряжении будут данные по ряду форм. Тогда же мы сможем установить точное положение Seymouria и Kotlassia, пока составляющих явственные семейства. По этим соображениям, я воздерживаюсь от уточненного диагноза Lanthanosuchus, общая диагностика которого ясна из описания.
Подведем краткий итог.
С развитием палеонтологических знаний все яснее вырисовывается существование обширной, переходной между классами амфибий и рептилий группы батрахозавров, несомненно ранее распространенной гораздо более широко. Батрахозавры явились непосредственными потомками форм, стоявших у начала расщепления обоих классов в самом начале карбона, и далее развивались, как самостоятельная группа, давая самые разнообразные эволюционные уклонения, в отдельных случаях обладавшие признаками высших рептилий.
Биологически батрахозавры несомненно также занимали промежуточное между Amphibia и Reptilia положение. Они обладали (частично) реберным дыханием, причем различные члены группы показывают разные степени его развития. Размножение батрахозавров, повидимому, представляло собою различные ступени перехода от метаморфоза амфибий к защищенным оболочкой и богатым желтком яйцам рептилий.
Среди современных амфибий мы знаем примеры весьма сильного сокращения метаморфоза и развития яиц не в воде, а во влажном субстрате почвы из растительных остатков и даже на деревьях. Болотные области развития батрахозавров как раз наиболее выгодны для постепенного перехода к безметаморфозному способу размножения.
Появление волокнистых оболочек вместе с укрупнением желточного мешка у яиц современных амфибий, развивающихся вне воды, и, с другой стороны, примитивные влагоемкие кожистые оболочки яиц таких современных рептилий, как гаттерии и ящерицы, дают нам основные вехи пути эволюции способа размножения батрахозавров.
Особняком стоит вопрос о кожных покровах, без существенных изменений которых немыслимо превращение амфибии во вполне наземную рептилию. Здесь мы пока не имеем данных даже для предположений. Некоторый свет на этот вопрос проливает открытие Ромером (Romer, 1941) отпечатков кожи Eryops — примитивного рахитомного стегоцефала из верхов карбона и низов перми США, синхроничного сеймурии. Кожа Eryops, принадлежащего к «наземным» стегоцефалам, защищена тончайшими костными чешуйками, очевидно, прикрытыми сверху тонким слоем эпидермиса, достигающим (ввиду крупной величины животного) наибольших размеров (около 10 мм по наибольшему диаметру овала). Подобное строение кожи вполне сходно с кожей современных безногих амфибий (Нуmnорhiоnа) и, в частности, с кожей Ichthyophis.
Если стегоцефалы обладали в древнейшие времена такой уже сравнительно защищенной кожей, то батрахозавры могли сделать еще один шаг вперед на пути к хорошо защищенной роговым покровом коже пресмыкающихся и приобрести затвердевший над костными чешуйками типа Eryops роговой слой, не нуждавшийся в смачивании, в то время как дополнительное усиление дыхания компенсировало ослабление газообмена через поверхность кожи.
Впрочем, все подобные сопоставления пока еще не могут считаться даже гипотезами.
Первой большой адаптивной радиацией наземных позвоночных была радиация стегоцефальных амфибий карбона. Она происходила в затопленных прибрежных и дельтовых лесах, представлявших идеальную среду для перехода от водной к наземной жизни. Батрахозавры, повидимому, обитали на краевых частях обширных болот, на переходе к сухой поверхности материка, и в своих реликтах (Kotlassia, Lanthanosuchus, Seymouria) остались теми же болотными формами. Поэтому батрахозавры отсутствуют в обычных захоронениях каменноугольных фаун, связанных исключительно с затопленными лесами. Адаптивная радиация батрахозавров, совершавшаяся в узком ареале их обитания в начале карбона, сменилась мощным развитием настоящих наземных позвоночных — рептилий, быстро освоивших все наземные экологические ниши, в связи с появлением растительноядных форм. Между этими двумя великими периодами формообразования наземных позвоночных затерялась адаптивная радиация батрахозавров той промежуточной ступени эволюции древнейших четвероногих, которая, собственно, и осуществила постепенный переход к полностью наземному существованию. Но недалеко то время, когда мы сможем восстановить ее в деталях. Много странных, биологически трудно объяснимых форм с ранним появлением отдельных признаков высокой организации — первых завоевателей границы водных и наземных биотопов материка — будет обнаружено дальнейшими раскопками.
Палеонтологический институт
Академии Наук СССР
Поступило
3 III 1946
ЛИТЕРАТУРА
Ефремов И. А. 1940. Предварительное описание новых форм пермской и триасовой фауны наземных позвоночных СССР. Тр. Палеонт. инст. АН СССР, 10. 2.
Ефремов И. А. 1941. Краткий обзор фауны пермских и триасовых Tetrapoda СССР. Советская Геология, 5.
Broili F. 1904. Permische Stegocephalen und Reptilien aus Texas. Palaeontographica, 51.
Broom R. 1913. On the structure and affinities of Bolosaurus. Bull. Amer. Mus. Nat. Hist., 32.
Bystrow A. P. 1944. Kotlassia prima Amalitzky. Bull. Geol. Soc. America, 55.
Cope E. D. 1896. Second contribution to the history of the Cotylosauria. Proc. Amer. Phil. Soc., 35.
Оlsоn E. C. 1936. The dorsal axial musculature of certain primitive Tetrapoda. Journ. Morph., 59.
Romer A. S. and Witter R. V. 1941. The skin of the Rhachitomous Amphibian Eryops. Amer. Journ. Sci.. 239, Novemb.
Sushkin P. P. 1925. On the representatives of the Seymouriamorpha, supposed primitive Reptiles, from the Upper Permian of Russia, and on their phylogenetic relations. Boston Soc. Nat. Hist. Occas.Papers, 5.
Sushkin P. P. 1927. On the modifications of the mandibular and hyoid arches and their relation to the braincase in the early Tetrapoda. Paläont. Ztschr., 8.
Watson D. M. S. 1918. On Seymouria, the most primitive known Reptile. Proc. Zool. Soc. London.
White T. E. 1939. Osteology of Seymouria baylorensis Broili. Bull. Mus. Compar. Zool., 85, 5.
J. A. EFREMOV. ON THE SUBCLASS BATRACHOSAURIA — AN INTERMEDIARY GROUP BETWEEN AMPHIBIANS AND REPTILES
SUMMARY
A new form of Lanthanosuchus watsoni, gen. et sp. nov. belonging to the family of Lanthanosuchidae is described. It has been discovered in the Isheyevski deposits (Middle Volga region) of the II zone of Upper Permian. A great ancient group is established, which is named Batrachosauria.
The indicated group is intermediary between amphibians and reptiles. Batrachosaurians represent a first radiation of terrestrial forms that took place in the second half of Carboniferous after the great radiation of Carboniferous amphibians.
Дата добавления: 2020-04-25; просмотров: 216; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!